| Ноябрь 2024 | | Пн | Вт | Ср | Чт | Пт | Сб | Вс |
|---|
| | | | | 1 | 2 | 3 | | 4 | 5 | 6 | 7 | 8 | 9 | 10 | | 11 | 12 | 13 | 14 | 15 | 16 | 17 | | 18 | 19 | 20 | 21 | 22 | 23 | 24 | | 25 | 26 | 27 | 28 | 29 | 30 | |  Календарь Календарь |
|
| Последние темы | » Передержка.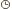 24/3/2019, 1:20 am автор Светланка » Болтаем обо всем на свете28/12/2017, 9:55 pm автор Пурш » Тех вопросы15/11/2017, 10:58 pm автор Злюсенький Admin » БРИТАНИЯ ТАЙГРЭСС ИЗ БУЛФАСТ`С + Зена Квин оф Варриорс+ Зикфрид Викториус. (Лиля +Боська + Зена+Зик).15/11/2017, 10:57 pm автор Злюсенький Admin » Кормить с подставки или без?15/11/2017, 10:55 pm автор Злюсенький Admin » ЗНАКОМИМСЯ И ПРЕДСТАВЛЯЕМСЯ)))12/5/2017, 2:07 pm автор Muftocka. » В Латвию вернулся опасный для кошек и собак вирус, схожий с бешенством2/2/2017, 4:03 pm автор Muftocka. » Гранд Канис Аниксия Мосдорм и Голден Бест Жордан Бои( Ника+Жорик+Ирина)12/1/2017, 2:47 pm автор Пурш » С НАСТУПАЮЩИМ НОВЫМ ГОДОМ!!1/1/2017, 10:01 pm автор Пурш |
| Кто сейчас на форуме | Сейчас посетителей на форуме: 11, из них зарегистрированных: 0, скрытых: 0 и гостей: 11
Нет
Больше всего посетителей (435) здесь было 25/10/2024, 11:30 am
|
|
| | Генетика собак |  |
| | | Автор | Сообщение |
|---|
Гость
Гость
| | Тема: Генетика собак 17/1/2014, 1:38 am | |
| ОСНОВЫ ГЕНЕТИКИ
СТРОЕНИЕ И ФУНКЦИЯ КЛЕТКИ
Любая собака, как и все другие животные, состоит из бесчисленного числа клеток , в основе которых лежит два основных компонента. Основную часть клетки занимает цитоплазма, состоящая преимущественно из белков и содержащая различные структуры. В центре цитоплазмы располагается более темный участок — ядро (см. рис. 1). Именно в нем находится основной генетический материал .
В ядре имеется ряд структур, называющихся хромосомами . Размер и форма хромосом (их тип) являются специфическим и постоянным фактором для каждого вида животных и растений (это закон постоянства формы хромосом). В любой соматической клетке живого организма содержится постоянное и характерное для него, как и для его вида, четное число хромосом (за исключением половых клеток) (закон постоянства числа хромосом). У собаки их 78, как и у всего семейства собачьих, включая волков и койотов; у шакала их 74, у человека — 46, у семейства кошачьих — 38, у лошадей — 64 и т. д. Следовательно, разные виды животных и растений различаются не только по числу хромосом, но и по природе наследственных признаков, содержащихся в них. Совокупность числа, размеров и особенностей строения хромосомного набора называется кариотипом (ядерным типом). Тщательное научное исследование показало, что независимо от размера и формы все хромосомы в соматической клетке расположены попарно (закон парности хромосом), то есть речь идет об их диплоидном наборе, в отличие от гаплоидного, имеющегося в половых клетках . Таким образом, правильнее говорить, что у собак не 78 хромосом, а 39 их пар.
На каждой хромосоме линейно располагаются гены . Для сравнения можно взять нитку бисера, в которой сама нитка — хромосома, а нанизанный на ней бисер — гены. Теоретически подобная аналогия неверна, так как гены — важнейшая составная часть хромосомы, а бисер существует сам по себе, вне зависимости от нитки. Но для наглядности такое сравнение вполне допустимо.
С химической точки зрения основу гена составляет ДНК (дезоксирибонуклеиновая кислота), благодаря которой передается огромная масса генетической информации и команд, на которых основывается наследственность. Ген — это единица наследственности. Как и хромосомы, гены располагаются попарно. Парные хромосомы, из которых одна происходит от материнского организма, а другая от отцовского, называют гомологичными. Как правило, гомологичные хромосомы данной пары морфологически неотличимы. Гены, занимающие в гомологичных хромосомах одно и то же место (локус, или локализация гена), отвечают за один и тот же признак/признаки и называются аллельнымиК Так как часть гомологичной хромосомы наследуется от отца, а другая от матери, то и аллельные гены наследуются точно так же.
Если аллеломорфная пара идентичная (в гомологичных хромосомах имеется пара одинаковых генов), то есть обе они отвечают за одно и то же проявление признака, то данную особь называют гомозиготной по этому признаку, а если каждый из этих аллелей отвечает за альтернативное (контрастное) проявление одного и того же признака, то гетерозиготной по нему.
Митотическое деление клетки
Щенок растет и увеличивается в размерах за счет деления соматических клеток, называемого митозом2. Митоз — непрямое деление соматической клетки, когда происходят сложные изменения в ее ядре и цитоплазме. После оплодотворения (ово- гамии) яйцеклетки сперматозоидом (слияния, или копуляции гамет) образуется зигота (ооциста) — новый организм, состоящий всего из одной клетки. Процесс роста и развития нового организма начинается с момента первого митотического деления этой клетки (материнской), когда из нее возникают две дочерние (точнее, сестринские), полностью схожие с ней клетки, и продолжается до смерти.
В процессе митоза происходит:
1) удвоение вещества хромосом;
2) изменение физического состояния и химической организации хромосом;
3) расхождение сестринских хромосом к полюсам клетки;
4) последующее деление цитоплазмы и полное восстановление двух ядер в новых клетках.
В митозе заложен жизненный цикл ядерных генов: удвоение, распределение и функционирование. Период между делениями клетки называется интерфазой, во время которой в ней протекают активные процессы жизнедеятельности и подготовка к следующему делению. Весь цикл изменений, происходящих в клетке
от одного ее деления до другого, называется митотическим циклом. Последний состоит из двух основных периодов — интерфазы и непосредственно митоза.
В результате митоза из одной клетки образуются две, имеющие идентичные хромосомы. Таким образом, митоз обеспечивает преемственность и постоянство числа и набора, то есть качественной специфичности хромосом в последовательных поколениях делящихся клеток (см. рис. 2).
Рис. 2. Митоз
1 — три пары хромосом; 2 — репликация хромосом с образованием сестринских хроматид, соединенных в области центромеры; 3 — у полюсов ядерной оболочки видны центросомы с астеральными лучами, идущими к центромер- ному участку хроматид, выстроенных по экватору, чтобы разъединить центромеры сестринских хроматид и развести их к разным полюсам; 4 — деспирали- зация хромосом, восстановление ядерной оболочки и образование клеточной перегородки с образованием идентичных материнской двух дочерних клеток с точно таким же, как в ней, числом хромосом
3
Мы не будем подробно останавливаться на всех фазах митоза. Скажу лишь одно, что в интерфазе, периоде между двумя последовательными делениями клетки, в ядре происходит репликация (аутодупликация, или самоудвоение) ДНК, а значит, и числа хромосом в клетке (образование сестринских хроматид, удерживаемых вместе центромерой, то есть тельцем, выполняющим функцию механического центра хромосомы), а также деспирализация последних. В метафазе, или центральной фазе деления ядра, хромосома, состоящая из двух хроматид, превращается в две дочерние хромосомы. В анафазе происходит деление и расхождение дочерних хромосом к полюсам клетки, то есть восстановление их
должного числа. В телофазе, конечной стадии деления клетки, хромосомы приобретают тот же вид, что и до начала деления, а количество ДНК в каждом дочернем ядре уменьшается вдвое по сравнению с предыдущими стадиями. Таким образом, обе дочерние клетки содержат одинаковые количества цитоплазмы и идентичные наборы хромосом и готовы пройти митоз.
Разумеется, постоянно делятся не все соматические клетки организма. В процессе эмбрионального развития происходит дифференциация органов и тканей, развивающихся по своему специфическому, генетически заложенному пути. Поэтому одни клетки превращаются в клетки мозга, другие — в клетки крови и т. д. Причем одни из них делятся постоянно, а другие лишь на определенной стадии развития или при необходимости, отвечая, например, за репаративные (восстановительные) процессы.
Мейотическое деление клеток
При половом размножении сходство потомков с родителями обеспечивается через половые клетки. Несмотря на свои ничтожные по сравнению с телом организма размеры, они несут в себе всю наследственную информацию, предопределяющую ход развития будущего организма.
Основой полового размножения является оплодотворение (сингамия), то есть слияние двух половых клеток.
Физиологическая специализация половых клеток обусловила особенности их морфологии и физиологии, из-за чего женские и мужские половые клетки значительно отличаются друг от друга. Яйцеклетка женского организма не только отвечает за передачу наследственной информации потомству, но и за питание эмбриона на ранних стадиях его развития. Мужская половая клетка, сперматозоид, этой функцией не обладает, а обеспечивает передачу наследственных свойств отцовского организма потомкам и стимулирует яйцеклетку к развитию.
В основе развития половых клеток лежит мейоз , или редукционное деление клеток. В него вступают незрелые половые клетки, достигшие определенной дифференциации. То есть он совершается в период образования гамет, или гаметогенеза.
Если бы каждая половая клетка имела диплоидный набор хромосом, заключающийся в соматической клетке, то число хромосом удваивалось бы в каждой последующей генерации. А так как кариотип у каждого вида животных постоянен, то число хромосом в гаметах должно быть гаплоидным. За уменьшение числа хромосом в гаметах (их редукцию) и отвечает мейоз.
Сущность мейоза заключается в том, что происходит два быстро следующих друг за другом деления ядра, а хромосомы при этом редуплицируются только один раз. Первое деление называется редукционным. При этом происходит уменьшение числа хромосом в ядрах вдвое за счет сближения гомологичных хромосом, называемого конъюгацией . При этом они, как правило, взаимно перекручиваются и состоят из двух соединенных центромерой хроматид. Во время конъюгации гомологичные хромосомы в местах перекручивания могут разрываться и обмениваться между собой гомологичными участками. В этот процесс, называемый кроссинговером , вовлекаются только две (любые)
хроматиды из четырех — по одной из каждой гомологичной хромосомы (см. рис. 3). Кроссинговер приводит к перекомбинации генетического материала в хромосомах, а это означает, что в гаметах будут не только хромосомы, идентичные родительским (не обменявшиеся участками), но и хромосомы с новой комбинацией материала двух гомологичных хромосом. Из каждой пары гомологичных хромосом материнской клетки в дочерние ядра попадает по одной, то есть гомологичные хромосомы, в отличие от негомологичных, которые комбинируются по-раз- ному, зависят друг от друга. Второе деление — обычное и называется эквационным, или уравнительным. При этом центромера каждой хромосомы делится и дочерние хромосомы (бывшие хроматиды) в равном, но уменьшенном вдвое количестве расходятся к противоположным полюсам клетки. В результате двух делений мейоза из одной диплоидной родительской клетки образуется четыре гаплоидных дочерних клетки, содержащие по 39 хромосом (по одной из пары гомологичных хромосом). После образования зиготы в ней окажется 78 хромосом из 39 гомологичных пар.
На рис. 4 вы видите четыре гаметы, в две из которых попали хромосомы А2В,С,, а в две другие — А,В2С2. Если под цифрой 1 мы обозначим хромосомы, попавшие в клетку от отцовского организма, а под цифрой 2 — от материнского, вы поймете, что их распределение в гаметах не имеет какой-либо закономерности, а носит совершенно случайный характер. Теоретически в каждую гамету попадает одинаковое количество хромосом матери и отца. Но не следует забывать о независимом поведении негомологичных хромосом и кроссинговере гомологичных хромосом во время мейоза, что сопровождается рекомбинацией генов1 и приводит к появлению мириад гамет, отличающихся от материнской клетки и между собой по набору хромосом, не говоря уже о том, что любая из гамет может участвовать в образовании зиготы.
Способы, которые позволяли бы нам контролировать при разведении собак случайное распределение хромосом и находящихся в них генов, неизвестны, что делает генетические прогнозы крайне сложными и увлекательными. Но из этого вовсе не следует, что мы должны быть фаталистами и считать плоды своей племенной работы чистой случайностью. Конечно, определенный элемент случайности при этом имеется, но
для достижения конкретных целей, стабилизации и улучшения породы заводчик обязан знать основы генетики, проводить тщательную селекцию, уметь подбирать и отбирать племенные пары, а также вести учет для статистической обработки данных. Чем строже он ведет отбор, чем больше знает о специфических генетических факторах, тем выше его шанс на успех.
Кроссинговер и сцепление
Согласно хромосомной теории гены в хромосомах расположены в линейном порядке, в определенных местах (локусах) и на определенном расстоянии друг от друга. Это доказывается с помощью явления кроссинговера (см. рис. 5). Гены, находящиеся в одной паре гомологичных хромосом, наследуются вместе, так что при гаметогенезе они попадут в одну гамету. Это гово-" рит о том, что они сцеплены1. Совместное наследование генов, ограничивающее их свободное комбинирование, называется сцепленным наследованием.
Группу сцепления образуют все гены, локализованные в одной хромосоме, так как каждый из них проявляет сцепление. Гены одной группы сцепления наследуются независимо от генов других групп сцепления. Число групп сцепления равно гаплоидному числу хромосом.
Частота мейотического перекреста между двумя генами, находящимися в одной хромосоме в определенных условиях среды, постоянна, что возможно лишь при линейном их расположении. Таким образом гены отцовской хромосомы могут переместиться в материнскую. Установлено, что сила сцепления между генами зависит от их расстояния друг от друга и обратно пропорциональна ему: чем дальше друг от друга расположены в хромосоме гены, тем меньше сила сцепления между ними и тем чаще происходит кроссинговер и наоборот. Она определяется путем вычисления процента кроссоверов (гамет с хромосомами, претерпевшими кроссинговер), возникших в результате перекреста хромосом по формуле:
число кроссоверов
Процент перекреста = х 100.
общее число потомков
Рис. 5. Кроссинговер
1 — гомологичные хромосомы (темная — отцовская); 2 — репликация хромосом с образованием двух сестринских хроматид; 3 — кроссинговер; 4 — окончательная комбинация хромосом
Чем эта сила больше, тем меньше сила сцепления.
Опытным путем было установлено, что перекрест между хромосомами может быть одиночный (в одной точке), двойной и множественный (в нескольких точках одновременно). Иногда перекрест, происшедший в одном участке хромосомы, способствует или препятствует наступлению другого перекреста в ближайших участках хромосомы, что называется интерференцией (вмешательством).
|
| | | | Гость
Гость
| | Тема: Re: Генетика собак 17/1/2014, 1:38 am | |
| В начале этой главы было сказано, что кариотип собаки представлен 39 парами хромосом, одна из которых отличается у кобелей и сук. Те хромосомы, по которым мужской и женский пол не различаются (38 пар), называются аутосомами. У кобелей имеется неодинаковая пара половых хромосом (XY), и при гаметогенезе у них образуются сперматозоиды двух сортов — половина с Х-хромосомой и половина с Y-хромосомой . У сук же обе половые хромосомы одинаковые (XX).
У птиц, некоторых рыб и насекомых гомогаметным полом являются самцы (ZZ), а гетерогаметным — самки (ZW), поэтому определение пола у них происходит не при оплодотворении, а еще при редукционном делении оогенеза (в зависимости от того, какую половую хромосому получит яйцеклетка).
У собак пол генотипически определяется в момент оплодотворения и зависит от того, каким сперматозоидом (несущим X- или Y-хромосому) будет оплодотворена яйцеклетка (всегда несущая Х-хромосому). Согласно мейозу, среди потомков получается 50% сук и 50% кобелей (соотношение 1:1) — см. рис. 6.
В том случае, когда гены находятся в аутосомах, реципрок- ные скрещивания дают одинаковые результаты, так как аутосо- мы у обоих полов одинаковые. Если же гены находятся в половых хромосомах, то наследование признаков, определяемое ими, будет зависеть от поведения половых хромосом в мейозе при образовании гамет и от особенностей этих хромосом. Y-хромо- сома наследственно инертна, так как не содержит генов. Это говорит о том, что в определении пола она не участвует, а гены мужского пола располагаются в аутосомах. Следовательно, гены, находящиеся в Х-хромосоме, аллелей в Y-хромосоме, как правило, не имеют. В результате даже рецессивные гены, находясь в Х-хромосоме и не имея аллели в Y-хромосоме, проявляются как у гомозиготы .
При повышении в генном балансе особи доли аутосом по отношению к количеству Х-хромосом у потомства усиливается проявление мужских половых признаков, а при соответствующем повышении в этом балансе доли Х-хромосом усиливается проявление женских признаков. В этом состоит суть балансовой теории определения пола и нарушение соотношения 1:1. Из этого следует, что пол особи определяется преобладанием генов одного пола над другим в этом балансе и что гены обоих полов в зиготе всегда присутствуют одновременно (в ней всегда имеются аутосомы с генами мужского пола и одна или две Х-хро- мосомы с генами женского пола в них), то есть любой организм бисексуален. В каждой зиготе одновременно заложены потенци-
х х
^ Сука ^^
Род
Гаметы
□ /в\ п
Зиготы
Сука Кобель Кобель
Сука
Рис. 6. Наследование пола
альные возможности развития как в женскую, так и в мужскую особь. Все зависит от характера взаимодействия женских и мужских генов в генном балансе, от соотношения силы их действия.
Так или иначе, особей мужского пола (XY) насчитывается больше, чем женского (XX). Но природа отсеивает их за счет ранней смертности и менее продолжительного срока жизни, так что, в конце концов, преобладают все-таки особи женского пола.
ВЗАИМОДЕИСТВИЕ ГЕНОВ - ПОЛНОЕ ДОМИНИРОВАНИЕ
Мы уже знаем, что гены построены из ДНК — вещества, являющегося хранилищем генетической информации. Нас, как заводчиков, интересует не столько происходящие с ней биохимические реакции, сколько их фенотипические проявления. Ген — участок молекулы ДНК, в котором содержится информация о первичной структуре одной цепи синтезируемого белка. Один ген — одна полипептидная цепь. Сами гены непосредственного участия в синтезе белка не принимают. Они служат молекулярной матрицей. Функция генов состоит в программировании
синтеза белков в клетке. Их первичным продуктом в ядре являются все виды РНК (рибонуклеиновая кислота), контролирующие синтез белков в клетке. Именно через эти белки гены контролируют синтез определенных продуктов в клетке, определяют ее функцию, влияя на течение онтогенеза организма.
Так, собака, например, наследует не ген подпала, а определенную цепочку полинуклеотидной цепи, входящую в состав ДНК (участок аминокислот, расположенных в строго определенном порядке и сочетании), которая в соответствующих условиях и в зависимости от происходящего на других участках ДНК приводит к появлению на шерсти пятен подпала. При любых нарушениях со стороны цепочки или в самой ДНК мы не получим этого окраса или даже самого щенка. Для простоты мы будем говорить о генах так, будто бы наследственность представляет собой однолинейный, а не сложный комплексный механизм.
Перейдем же к изучению различных типов взаимодействий аллельных генов.
При полном доминировании, отвечающем первому закону Менделя (единообразия гибридов первого поколения), один ген аллеломорфной пары подавляет проявление другого. В этом случае он называется доминантным (от лат. dominantis — преобладающий, господствующий), а тот, что не смог проявиться, оставшись в скрытом состоянии, — рецессивным (от лат. recessus — отступление). Из этого следует, что доминантный ген проявляется независимо от того, находится ли он в обеих гаметах, образующих зиготу, то есть в гомозиготном состоянии, или в одной из них, то есть в гетерозиготном состоянии, а рецессивный — лишь в гомозиготном, то есть если он имеется и в материнской, и в отцовской гаметах.
Совокупность всех генов организма, взаимодействующих между собой и условиями среды, называется генотипом (от греч. genos — род и typos — отпечаток, образец, то есть генный или наследственный тип организма), а их внешнее проявление в совокупности с индивидуальным развитием особи — фенотипом (от греч. phaino — являю и typos, или внешний вид организма).
Прежде чем перейти к дальнейшему описанию, скажу несколько слов об обозначениях, введенных в генетике Г. Менделем, У. Бэтсоном и Сондерсом. Скрещивание обозначается знаком умножения — х, доминантный признак и контролирующий его наследственный фактор — прописной буквой латинского алфавита (например, А), а рецессивный — строчной (а), причем аллельные гены обозначаются одной и той же буквой (АА, Аа или аа); знак доминирования — >; при написании схемы скрещивания на первое место ставят материнский организм, обозначающийся зеркалом Венеры —9, а на второе — отцовский, обозначающийся щитом и копьем Марса — с?; исходные родительские формы обозначают латинской буквой — Р (от лат. parentale — родители), а организмы, полученные от них, — F (от лат. filii — дети), причем с цифровым подстрочным индексом, означающим гибридное поколение и его порядковый номер, например F,, F2, F3 и т. д.
Для примера рассмотрим скрещивание суки, гомозиготной по доминантному черному окрасу — ВВ (от англ. black — черный), с кобелем рецессивного печеночного (коричневого) окраса — bb .
Р
Генотип В В х bb
Фенотип черный печеночный
Гаметы В В b b
F.
Генотип Bb
Фенотип черный
В данном случае мы говорим о моногибридном скрещивании, так как родительские организмы отличаются по одной лишь паре контрастных альтернативных признаков. Как видно из примера, все гибриды первого поколения имеют черный окрас шерсти. Преобладание у гибрида первого поколения признака одного из родителей Мендель назвал доминированием. Поскольку все гибриды F, одинаковые (черные), закон доминирования чаще называют законом единообразия гибридов первого поколения (первый закон Менделя).
При скрещивании гибридов F, между собой во втором поколении появятся особи как черного, так и коричневого окрасов, то есть с признаками обоих родителей. Это подчиняется второму закону Менделя — закону расщепления признака .
F,
Генотип Bb х Bb
Фенотип черный черный
Гаметы В b В b
f2
Генотип BB 2Bb bb Расщепление: 1:2:1
Фенотип черный черный печеночный Расщепление: 3:1
Из того факта, что рецессивный признак (печеночный окрас), отсутствовавший у гибридов первого поколения Fp вновь появляется у '/4 гибридов F2, вытекает, что наследуются не сами признаки, а гены, их определяющие. При образовании половых клеток в каждую гамету попадает только один из каждой пары соответствующих генов (один доминирующий над другим), что отвечает третьему закону Т.Х. Моргана — закону чистоты гамет.
В качестве непременного элемента в систему гибридологического анализа входит скрещивание гибрида F, с рецессивной родительской формой — анализирующее скрещивание, или бэк- кросс (от англ. back — назад, обратно и cross — скрещивать, пересекать), подтверждающее закон чистоты гамет. Потомство от такого скрещивания обозначают Fb.
bb
Bb черный В b
Генотип Фенотип Гаметы
печеночный b b
bb
Генотип Фенотип
Bb черный
печеночный
Расщепление: 1:1 Расщепление: 1:1
Как видно из схемы, расщепление по генотипу и фенотипу совпадает. Это скрещивание позволяет анализировать генотип гибрида, так как рецессивная родительская форма дает лишь один сорт гамет с рецессивным аллелем Ь, что позволяет проявиться в фенотипе потомков Fb любому аллелю, пришедшему с гаметами гетерозиготного родителя.
В некоторых случаях используется реципрокное скрещивание (от лат. recipricus — взаимный, обратный) — взаимно прямое и обратное скрещивание, при котором особи одной породы (например, А и В) в одном случае используются в качестве материнской, а в другом — в качестве отцовской форм (то есть 9А хс?5 и x<jl4). Один из этих вариантов скрещивания условно называют прямым, а другой — обратным.
Ряд комбинаций двух этих аллелей показан в табл. 1.
ТАБЛИЦА 1 процентное соотношение Fl/Фенотип
Р/Фенотип ВВ ВЬ ьь
черный черный печеночный
ВВ х ВВ 100 — —
ч п
ВВ х ВЬ 50 50 —
ч ч
ВВ х bb — 100 —
ч п
ВЬ х ВЬ 25 50 25
ч ч
ВЬ х bb — 50 50
ч п
bb х bb — — 100
п п
Эти данные будут статистически достоверными лишь при анализе не менее ста щенков. В противном случае вам трудно будет судить о генотипе двух родителей черного окраса, так как у вас может не оказаться в помете щенков печеночного окраса, несмотря на то, что один из них или оба гетерозиготные. А вот рождение из-под них хотя бы одного щенка печеночного окраса четко указывает на их гетерозиготность.
Кроме анализирующего скрещивания, часто пользуются анализом родословных, при составлении которых применяют специальные символы. Персона, с которой начинают составление родословной, называется пробандом. Если она спускается к потомству, ее называют генеалогическим древом, а если восходит к предкам — таблицей предков. При анализе родословных можно определить, наследуется ли интересующий нас признак, а если да, то как — по доминантному или по рецессивному типу. Если вам требуется определить количественные отношения при расщеплении, анализируют несколько сходных родословных.
Изучение наследования рецессивных признаков более трудное, так как в этом случае возможен перескок признака через генерации, то есть создается впечатление его неожиданного появления (см. рис. 7).
66Ь
йсхк^п
ййтоббб^то йй-гоб
*6Ъ
DrO
□тбОтП
DjO
Й15В
Пгб Сюббуй
ОтО
151 й
Рис. 7.
1. Пример родословных с доминантными аномалиями
2. Пример родословных с рецессивными признаками
3. Пример родословной с семейственной рецессивной аномалией (эпилепсия)
О пол не выяснен ЯШ обладатель изучаемого признака О гетерозиготный носитель изучаемого рецессивного гена д рано умер О-а вязка
ОО тесный инбридинг (отец
с дочерью, мать с сыном) ОтО родители
Наиболее употребительные символы, принятые при составлении родословных □ кобель О сука
©Шдети и порядок их рождения
Наследование признаков при скрещивании подчиняется статистическим закономерностям и законам вероятности, если число потомков достаточно велико.
ТАБЛИЦА 2
Результаты, ожидаемые при разных видах скрещивания
Вид скрещивания 1 2 3 4 5 6
п* 2" (2")2 2" 3" (3:1)"
Моногибридное 1 2 4 2 3 3:1
Дигибридное 2 4 16 4 9 9:3:3:1
Тригибридное 3 8 64 8 27 27:9:9:9:3:3:3:1
1 — число контрастных альтернативных признаков, по которым отличаются скрещиваемые особи (число пар генов); 2 — число типов гамет, образующихся у гетерозигот; 3 — число возможных комбинаций гамет в F,; 4 — число классов разных фенотипов в F2; 5 — число разных генотипов в F2; 6 — число фенотипов в F2.
* — число пар контрастных альтернативных признаков.
|
| | | | Гость
Гость
| | Тема: Re: Генетика собак 17/1/2014, 1:41 am | |
| Доминирование не всегда бывает полным из-за промежуточного наследования, при котором гетерозиготные потомки по степени выраженности признака не похожи ни на одного из родителей, а занимают между ними как бы промежуточное положение. В этом случае видоизменяется и расщепление во втором поколении по фенотипу (вместо 3:1 оно будет 1:2:1).
Для примера возьмем две чистые линии (гомозиготных родителей) — мать с висячими ушами — НН, а отца — со стоячими — hh.
Р
Генотип НН х hh
Фенотип висячие стоячие
Гаметы Н Н h h
Fi
Генотип Hh
Фенотип полустоячие
F.
Генотип Hh х Hh
Фенотип полустоячие полустоячие
Гаметы Н h Н h
f2
Генотип НН 2Hh hh Расщепление: 1:2:1
Фенотип висячие полустоячие стоячие Расщепление: 1:2:1
Как видно из схемы, при неполном доминировании, в отличие от полного, во втором поколении расщепление по фенотипу и генотипу совпадает.
Одна из наиболее частых причин, вызывающая отклонения от менделевских количественных закономерностей расщепления, — неодинаковая жизнеспособность зигот разных генотипов (часто из-за наличия летальных генов).
Подобный пример можно привести и на основе наследования серии генов агути у длинношерстной колли. Нас интересуют в данном случае два аллеля — а?, отвечающего за чисто- соболий окрас, и а1 — за черно-подпалый (биколор). Остальные гены отвечают за появление при этих аллелях белых пятен на шерсти, характерных для колли, поэтому мы проигнорируем их.
Итак, возьмем чистые линии и повяжем суку оленьего окраса — ауау с черно-подпалым кобелем аха\
Р
Генотип ауау х а'а1
Фенотип олень ч/п
Гаметы ау а1
F,
Генотип ауа'
Фенотип черный с собольим налетом (ч/с)
х
ч/с
а" а1
Генотип а"а' Фенотип ч/с Гаметы а" а1
Генотип ауау 2ауа"
Фенотип олень ч/с
а'а" Расщепление: 1:2:1 ч/п Расщепление: 1:2:1
Предсказывать результат вязок в данном случае будет легче, если вы воспользуетесь таблицей 1, заменив в ней ген В на ау, a b — на а1.
ЭПИСТАЗ
До этого мы рассматривали с вами взаимодействие гомологичных генов, расположенных в гомологичных участках хромосом (в одном и том же локусе). Но взаимодействовать могут и гены, лежащие в разных локусах. Явление, когда ген одной аллельной пары подавляет действие гена другой аллельной пары (А > В или В > А), называется эпистазом, или супрессией. Гены-подавители называются ингибиторами, или супрессорами и обозначаются буквами I, S или Sn, а подавляемый — гипостатическим. В случае доминантного эпистаза ингибитор доминантный, а при рецессивном — рецессивный.
В качестве примера возьмем чистые линии и повяжем суку серого окраса — AAbb с черным кобелем — ааВВ (речь идет о ди- гибридном скрещивании, поэтому скрещивание гибридов первого поколения между собой расписывается в решетке Р.К. Пённе- та, в которой гаметы суки пишутся по горизонтали, а кобеля — по вертикали).
Р
Генотип AAbb
ааВВ
х
черный
а В
Фенотип серый Гаметы A b
F,
Генотип Фенотип
АаВЬ серый
АВ Ав аВ ав
АВ ААВВ ААВЬ АаВВ АаВЬ
серый серый серый серый
Ab ААВЬ AAbb АаВЬ Aabb
серый серый серый серый
аВ АаВВ АаВЬ ааВВ ааВЬ
серый серый черный черный
ab АаВЬ Aabb ааВЬ aabb
серый серый черный рыжий
12АЬ + ЪаВ + lab = 12 серых + 3 черных + 1 рыжий, или 12:3:1
Из схемы видно, что ген а > В и а > Ъ.
Явление эпистаза напоминает первый закон Менделя. Разница заключается в том, что при последнем доминантный ген подавляет действие рецессивного гена, принадлежащего к той же аллеломорфной паре, а при эпистазе один доминантный ген подавляет действие другого доминантного гена, принадлежащего к другой аллеломорфной паре. Следовательно, явление эпистаза выражается в неаллельном подавлении действия доминантного гена другим доминантным геном, принадлежащим к другой аллеломорфной паре.
Как видно из дигибридного скрещивания (когда родители отличаются по двум парам контрастных альтернативных признаков), негомологичные хромосомы, в отличие от гомологичных, распределяются в гаметах при гаметогенезе независимо друг от друга. Независимое наследование признаков отвечает четвертому закону Менделя, а образование новых комбинаций признаков при этом называется комбинативной изменчивостью.
В случае доминантного эпистаза наблюдается два типа расщепления среди гибридов второго поколения — 12:3:1 и 13:3, причем последнее происходит тогда, когда рецессивная форма фенотипически сходна с формой, имеющей эпистатический ген (а = В).
Давайте рассмотрим пример рецессивного эпистаза. Окрас доберманов обусловливается рядом совместного действия различ
ных генов, ведущими из которых являются уже рассмотренные нами гены В и Ь, а также ген D, отвечающий за глубину окраса, и ослабляющий его — ген d (от англ. dilution — разбавлять, разжижать). При наличии в генотипе аллельной пары генов dd, черно- подпалая собака (ч/п) будет голубо-подпалой (г/п), а красно-под- палая (к/п) — олене-подпалой (о/п) или даже изабелловой.
Р
Генотип Фенотип Гаметы
F,
Генотип Фенотип
> < BD Bd bD bd
BD BBDD BBDd BbDD BbDd
ч/п ч/п ч/п ч/п
Bd BbDd BBdd BbDd Bbdd
ч/п г/п ч/п г/п
bD BbDD BbDd bbDD bbDd
ч/п ч/п к/п к/п
bd BbDd Bbdd bbDd bbdd
ч/п г/п к/п o/n
BbDd ч/п
F,
Генотип Фенотип
9BD + 3Bd + 3bD + lbd = 9 ч/п + 3 г/п + 3 к/п + 1 о/п, или 9:3:3:1
bbDd к/п bD bd
BBDd ч/п BD Bd
BbDD 2BbDd Bbdd Расщепление: 1:2:1 ч/п ч/п г/п Расщепление: 3:1
Нетрудно заметить, что ген В > b при наличии гена D. Но если рецессивный ген d находится в гомозиготном состоянии (dd), он эпистатичен по отношению к генам черного и коричневого окрасов. Идентифицировать генотип по фенотипу можно лишь у добермана оленьего окраса.
Скрещивание доберманов с различным генотипом
ТАБЛИЦА 3
Генотип родителей Фенотип потомков (в
Черный Красный Голубой Олений
BBDD BBDd BbDD BbDd bbDD bbDd BBdd Bbdd bbdd
BBDD х BBDD 100 — — — — — — — —
х BBDd 50 50 — — — — — —
х BbDD 50 — 50 — — — — — —
x BbDd 25 25 25 25 — — — — —
x bbDD — — 100 — — — — — —
x bbDd — — 50 50 — — — — —
x BBdd — 100 — — — — — — —
x Bbdd — 50 — 50 — — — — —
x bbdd — — — 100 — — — — —
BBDd x BBDd 25 50 — — — — —
x BbDD 25 25 25 25 — — — — —
x BbDd 12'/, 25 12'/, 25 — — 12'/, 12'/, —
x bbDD — — 50 50 — — — — —
x bbDd — — 25 50 — — — 25 —
x BBdd — 50 — — — — 50 — —
x Bbdd — 25 — 25 — — 25 25 —
x bbdd — — — 50 — — — 50 —
BbDD x BbDD 25 — 50 — 25 — — — —
x BbDd 12'/, 12'/, 25 25 12'/, 12'/, — — —
x bbDD — — 50 — 50 — — — —
x bbDd — — 25 25 25 25 — — —
x BBdd — 50 — 50 — — — — —
x Bbdd — 25 — 50 — 25 — — —
x bbdd — — — 50 — 50 — — —
BbDd x BbDd 6'/4 12'/, 12'/, 25 6'/4 12'/, 6'/4 12'/, 6'/4
x bbDD — — 25 25 25 25 — — —
x bbDd — — 12'/, 25 12'/, 25 12'/, — 12'/,
x BBdd — 25 — 25 — — 25 25 —
x Bbdd — 12'/, — 25 — 12'/, 12'/, 25 12'/,
x bbdd — — — 25 — 25 — 25 25
bbDD x bbDD — — — — 100 — — — —
x bbDd — — — — 50 50 — — —
x BBdd — — — 100 — — — — —
x Bbdd 50 — — — 50 — — —
x bbdd — — — - - 100 - - -
bbDd x bbDd — — — — 25 50 — — 25
x BBdd — — — 50 — — — 50 —
x Bbdd — — — 25 — 25 — 25 25
x bbdd — — — — — 50 — - 50
BBdd x BBdd — — — — — — 100 — —
x Bbdd 50 50 —
x bbdd 100 —
Bbdd x Bbdd 25 50 25
x bbdd 50 50
bbdd x bbdd 100
НАСЛЕДОВАНИЕ ПРИЗНАКОВ, СЦЕПЛЕННЫХ С ПОЛОМ
Мы уже говорили, что особи мужского пола несут половые хромосомы XY, а женского — XX. Гены, находящиеся в половых Х-хромосомах, называются сцепленными с полом. В Х-хромосо- ме имеется участок, для которого в Y-хромосоме нет гомолога. Поэтому у особей мужского пола признаки, определяемые генами этого участка, проявляются даже в том случае, если они рецессивные. Эта особая форма позволяет объяснить наследование признаков, сцепленных с полом.
Сук, гетерозиготных по любому из сцепленных с полом признаков, называют носителями соответствующего рецессивного гена. Они фенотипически нормальные, но половина их гамет несет рецессивный ген.
Я уже отмечал, что соотношение полов у всех животных 1:1, что напоминает расщепление при моногибридном скрещивании, когда одна из скрещиваемых форм является гетерозиготной {Аа), а вторая — гомозиготной по рецессивным аллелям (аа).
Наследование признаков, сцепленных с полом, подчиняется закону Т.Х. Моргана и имеет отклонение от второго закона Менделя у гибридов второго поколения, если проводить анализ потомства без учета пола щенков.
Наследование от матери к сыновьям, а от отца — к дочерям называется крисс-кросс-наследованием, причина которого состоит в различии половых хромосом суки и кобеля.
Скрещивание гетерозиготной суки с гомозиготным кобелем из первого поколения (F,), по существу, является анализирующим, причем сам кобель представляет собой генетический анализатор, так как образует гаметы и с Х-хромосомой, несущей
Рис. 8. Родословная с аномалией, передающейся через Y-хромосому и встречающейся исключительно у кобелей
рецессивный аллель (ее получат суки F2), и с Y-хромосомой (ее получат кобели F2), что наглядно видно на рис. 8.
Y-хромосома меньше Х-хромосомы, поэтому содержащие ее сперматозоиды легче тех, которые содержат Х-хромосому, и могут плыть несколько быстрее, поэтому кобелей рождается больше, чем сук. Но смертность кобелей, как я уже подчеркивал, выше, чем у сук, благодаря чему поддерживается равновесие полов в окружающей среде.
У собак сцепленное наследование связано с присутствием некоторых генов в половой Х-хромосоме: гены крипторхизма — с, гемофилии А и Б — Л, катаракты — Cat, дисплазии бедра — HD, бесшерстности — Нг, дисплазии конъюнктивы — Cd и др. К заболеваниям, сцепленным с полом, относятся сахарный диабет, врожденная эпилепсия, атрофия поджелудочной железы, аутоиммунные заболевания, врожденный лимфоотек — Ly, окрас Мерль — Ми ряд других.
Для написания генотипа с генами, сцепленными с полом,
АВ А
используют написание ——, несцепленных с полом — — .
ab а
Сцепленное наследование признаков выявляется по преобладанию в потомстве гетерозиготы форм с родительскими сочетаниями признаков (по сравнению со свободным и независимым наследованием) и уменьшенному количеству форм с новыми сочетаниями признаков — рекомбинантов (при независимом наследовании признаков в потомстве от анализирующего скрещивания дигетерозиготы организмы с родительскими сочетаниями признаков и рекомбинанты встречаются с равной частотой — 1:1).
Г~
XY
XX
нООи
Рис. 9. Наследование гемофилии А и Б, рецессивного заболевания, сцепленного с Х-хромосомой, через суку-кондуктора п — здоровый с — кондуктор h — больной гемофилией
Поскольку пол потомков зависит преимущественно от кобеля, следовательно, любой ген Y-хромосомы будет передаваться от отца к сыновьям, но не к дочерям (ведь у них нет Y-хромосомы). По сравнению с этим суки передают сыновьям и дочерям Х-хро- мосому, правда не всегда с идентичными генами.
Я уже подчеркивал, что Y-хромосома является генетически инертной и отвечающей за проявление вторичных мужских половых признаков. В отличие от Х-хромосомы, в ней почти или вовсе нет генов. У человека Х-хромосома несет гены, вызывающие у мужчин дальтонизм и гемофилию, у кошек — черепаховый окрас, а у собак — гемофилию. При этом женщины и особи женского пола являются переносчиками (кондукторами) этих генов без фенотипического проявления заболеваний.
Известно, что все сцепленные с полом гены — рецессивные. На рис. 9 показано наследование гемофилии потомками мужского пола — от суки — носительницы (кондуктора) гена этой болезни и здорового кобеля. Как видите, одна половина дочерей и сыновей — здоровая, а вторая — суки — кондукторы аномалии и больные кобели. Несмотря на то, что они несут всего один рецессивный аллель Л, они страдают гемофилией, так как Y-хромосома генетически пустая.
При наличии подобных гемофилии летальных генов большинство кобелей погибает на ранних этапах жизни, так что они
Рис. 10. Наследование гемофилии А и Б через достигшего фертильности больного кобеля п — здоровый с — кондуктор h — больной гемофилией
XX
Hwh
XY
с
XX
XY
XX
XY
Н0°
№
ьцун
с
hWh
h
Рис. 11. Наследование гемофилии А и Б через достигшего фертильности больного кобеля и суку—кондуктора заболевания п — здоровый с — кондуктор h — больной гемофилией
не передают их потомству, благодаря чему не появляются и суки — кондукторы аномалий. В противном случае, если больному кобелю все же удастся дожить до зрелого возраста и дать потомство (что возможно при наличии полулетальных генов), он даст часть больных кобелей и сук — носительниц заболевания.
При скрещивании здоровой суки с больным кобелем все кобели в потомстве будут здоровыми, а суки — кондукторами аномалии (см. рис. 10). При скрещивании же больного кобеля с сукой — носительницей заболевания, половина кобелей в потомстве будет нормальной, половина — больной, половина сук — больной, а половина — кондукторами аномалии (см. рис. 11).
|
| | | | Гость
Гость
| | Тема: Re: Генетика собак 17/1/2014, 1:43 am | |
| Некоторые признаки, называемые зависимыми от пола, определяются генами, находящимися не в Х-хромосомах, а в одной из пар аутосом. Однако проявление таких признаков зависит от пола животного (независимо от сцепления с полом), что обычно связано с действием мужских или женских половых гормонов (в рецессивном состоянии признак отсутствует независимо от пола).
Типичными примерами являются все вторичные половые признаки — комолость у некоторых пород овец, раннее облысение у людей, яйценоскость кур и наследование окраса у эр- ширских молочных коров, как показано в таблице 4.
ТАБЛИЦА 4
Зависимость окраса эрширских молочных коров от пола
Генотип Фенотип
Быки Коровы
ММ Махагон Махагон
Mm Махагон Красный
mm Красный Красный
Некоторые признаки, ограниченные полом, проявляются только у самок, как, например, молочность, или у самцов, как крипторхизм, хотя носителями их могут быть и те и другие (так как они полигенные, а контролирующие их гены расположены в любых парах аутосом). Это осложняет генетические исследования и в некоторых случаях означает, что при селекции следует руководствоваться качеством потомства в целом, а не отдельно взятого животного, так как по одному полу невозможно сделать правильные выводы.
Некоторые признаки, зависящие от пола, проявляются и у самцов, и у самок, но у одних из них они встречаются чаще, а у других — реже. Так, заячья губа и подагра, встречающиеся у людей, чаще всего наблюдаются все-таки у мужчин, а вот spina bifida (расщелина позвоночника — врожденная аномалия, характеризующаяся неполным закрытием позвоночного канала из-за отсутствия дуги нескольких позвонков, главным образом в поясничной области) и дисплазия тазобедренного сустава — у женщин.
Наследование ограниченных полом признаков осуществляется в соответствии с законами Менделя.
НЕПОЛНАЯ ПЕНЕТРАНТНОСТЬ
При обсуждении доминантных и рецессивных аллелей предполагалось, что конкретный признак проявляется у животного в присутствии одного-единственного доминантного аллеля. Рецессивный же аллель для его проявления должен быть в гомозиготном состоянии. В этом случае мы имеем дело с так называемой полной (абсолютной) пенетрантностью {от лат. penetrare — проникновение). В некоторых же случаях, чаще всего у гетерозиготных особей (Аа), доминантный ген А в 75% случаев может вести себя как доминантный, а в 25% случаев — как рецессивный. В этом случае речь идет о неполной, а 75%-ной пенетран- тности. Причиной тому чаще всего служит полигенное наследование признака.
При неполной пенетрантности заводчик сталкивается с определенными трудностями, ибо на первый взгляд результаты вязок не соответствуют законам Менделя, что связано с недостаточным количеством потомства для его анализа.
ФЕНОКОПИИ
Заводчики часто ошибочно полагают, что наследственность зависит только от генотипа, забывая о факторах индивидуального развития организма и влияния внешней среды. Так, травма суки во время щенности может значительно повлиять на развитие эмбриона, но не на его генотип, в отличие от внешних воздействий, способных вызвать мутации у плода. Особь с измененным генотипом называется фенокопией.
Пожалуй, самым трагичным примером фенокопий у человека можно назвать случаи, связанные с применением беременными женщинами в 60-х годах талидомида — транквилизатора с тератогенным действием, приводящего к фокомелии или ахе- роподии у новорожденных (укорочению или отсутствию конечностей).
Примером фенокопии служат и мягкие хрящи у собак со стоячими в норме ушами. Конечно, их можно поставить за счет фиксации ушных раковин, но такие собаки стойко будут передавать полустоячие уши своему потомству.
ВАРИАЦИИ ЧИСЛА ПОЛОВЫХ ХРОМОСОМ
Я уже писал, что в норме кариотип собак включает 78 хромосом, образующих 39 пар, включая одну половую: XX — у сук и XY — у кобелей. При изучении генного баланса, генетики обнаружили, что, даже если он и нормальный, в нем все равно встречаются отклонения. Вариации числа половых хромосом могут быть связаны с целым набором хромосом (эуплоидия) или с одной хромосомой (анэуплоидия).
Пожалуй, самым известным примером такого явления служит синдром Дауна у человека, когда в двадцать первой паре хромосом (в норме их 23) появляется лишняя хромосома. Это происходит на определенной стадии формирования гамет и чаще наблюдается среди старородящих, как правило сорокалетних матерей.
Возможно, подобные хромосомные аберрации происходят чаще, чем нам кажется, но мы не так часто видим их проявле-
Рис. 12. Типы нехваток хромосом
1 — концевая (терминальная, или дефишиенси); 2 — две концевые нехватки, приводящие к образованию кольца; 3 — внутрихромосомная нехватка (делеция)
ние из-за гибели гамет или ранней смертности плода. Болезнь Дауна (одна из форм олигофрении) — один из тех случаев, когда аберрация не настолько серьезная с генетической точки зрения, чтобы вызвать раннюю смерть эмбриона. А так как подобные аномалии встречаются в собаководстве чрезвычайно редко, мало кто из ученых придает им большое значение.
Второй тип — дупликация (удвоение) хромосом. В этом случае одновременно с возникновением нехватки в результате разрыва хромосомы и потери участка внутри одной хромосомы могут накапливаться идентичные участки, несущие одни и те же гены. При дупликации происходит фенотипическое изменение признаков.
Если в норме каждый участок хромосомы представлен одной дозой гена, то при удвоении или утроении несущего эти гены участка доза гена увеличивается в два или три раза. То есть у сук имеется не диплоидный набор (2Х:2А, то есть половой индекс, или соотношение между Х-хромосомами и аутосомами, равен единице), а три половые хромосомы — XXX, из-за чего они называются сверхсуками (они могут быть диплоидными — ЗХ:2А с
половым индексом 1,5 или триплоидными — ЗХ:ЗА). У таких сук наблюдается нарушение астральных циклов (как правило, они стерильные), а если и нет, то при их вязке с нормальными кобелями рождаются интерсексы (2Х:ЗА с половым индексом 0,66) мужского или женского типов — гермафродиты. У кобелей комплекс XXY (синдром Клайфельтера) также сопровождается явлениями инфантилизма из-за недоразвития семенников и азооспермии. Лишняя Х-хромосома у котов обусловливает появление в помете самок черепахового окраса, реже самцов с половым набором XXY, страдающих стерилитетом.
Животные с лишней половой хромосомой, как правило, стерильные. Типичный пример — мул (гибрид кобылы, имеющей 64 хромосомы, и осла, у которого их 62), кариотип которого включает 63 хромосомы. Реже у сук имеется всего лишь одна половая хромосома — Х0, что проявляется в синдроме Шерешевского-Тёрнера (с явлениями инфантилизма и отсутствием яичников). Кобели с набором XY:3A (половой индекс 0,33), вместо XY:2A (половой индекс равен 0,5), чрезмерно мускулистые, грубого сложения, с массивным костяком и называются сверхкобелями. Они также бесплодны.
Иногда у кобелей бывает лишняя Y-хромосома, в этом случае у них, как и у мужчин с набором XYY, отмечается неоправданная и неуправляемая агрессия.
НЕКОТОРЫЕ БИОХИМИЧЕСКИЕ АСПЕКТЫ
В 1977 году М. Бёрнс [141], обсуждая мою книгу «Немецкая овчарка», опубликованную в 1976 году, высказала мнение, что я придерживаюсь несколько старомодного подхода к основам генетики. Это критическое замечание остается справедливым и по отношению к данной книге. С моей точки зрения, заводчики лучше усваивают генетику, объясняемую с точки зрения доминантных и рецессивных признаков, чем в свете более современных биохимических процессов. Однако замечание М. Бёрнс вполне справедливо, и для упрощенного объяснения действия генов я все-таки коротко поясню некоторые биохимические аспекты.
До этого я привел вам в качестве аналогии нитку бисера для иллюстрации понятия генов и хромосом. В строгом смысле она некорректна, ибо гены — не изолированные объекты, а составляющие части хромосомы. Вся информация, необходимая для определения наблюдаемых нами в отдельной особи
Один нуклеотид-
Один нуклеотид
Сахаро фосфат- Пары комплементарных " (Сахарофосфатный остов t оснований ный остов
Одна полинуклео- н-связь Одна полинуклео- тидная цепь . . тидная цепь
Рис. 14. ДНК (схематическое изображение развернутых цепей) А — аденин I
I Сахарофосфатный '.ОСТОВ
Две антипараллельные цепи
2 нм
Рис. 13. Схематическое изображение структуры ДНК. На один полный оборот спирали приходится 10 пар оснований
3,4 км
Основания-
кольца
Г — гуанин I Т — тимин ] Ц — цитозин I
одно кольцо
О фосфа°тДНЫе СВЯЗИ \ / Дезоксирибоза (сахар)
признаков, содержится в структуре ДНК, из которой состоят гены. Сама ДНК — полимер, менее сложный по сравнению с белками, состоящий из сахара (дезоксирибозы), остатков молекул фосфорной кислоты и четырех азотистых оснований (пу- риновых — аденина, гуанина и пиримидиновых — цитозина и тимина). Молекула нуклеиновой кислоты представляет собой цепочку мономеров, специфических для этих кислот — нукле- отидов. Строение определенной молекулы нуклеиновой кислоты характеризуется определенной последовательностью нукле- отидов. Каждый нуклеотид состоит из трех разных элементов: азотистого основания, углеводного компонента и фосфорной кислоты.
Для нуклеотидов ДНК характерна способность образовывать самые длинные молекулы из всех известных. Вероятно, каждый ген в хромосоме представляет собой участок ДНК, содержащий сотни или тысячи нуклеотидов.
В 1953 году американский биохимик Джеймс Дьюи Уотсон и английский биофизик и генетик Фрэнсис Харри Комптон Крик выдвинули гипотезу, что молекула ДНК представляет собой двойную спираль, напоминающую лестницу со ступеньками из азотистых оснований, соединенных строго специфически (аденин с тимином, а гуанин с цитозином), перилами которой служат остатки фосфорной кислоты и пентозного сахара. Они же создали модель молекулы ДНК, а в 1962 году они вместе с английским биофизиком Морисом Уилкинсом впервые получили высококачественные рентгенограммы молекулы ДНК (см. рис. 13). Цепочки полинуклеотидов этой спирально завитой двойной молекулы расположены так, что сахар и фосфорные группы лежат снаружи молекул, а основания, соединенные между собой водородными связями, — внутри молекул (см. рис. 14).
В отдельной цепочке из двадцати аминокислот хранится информация, отвечающая за синтез определенного белка, считываемая информационной, или матричной, РНК и переносимая транспортной РНК к рибосомной РНК. В определенных условиях двойная цепь ДНК разрывается на месте водородных связей и возле каждой цепи строится новая комплементарная цепь с помощью фермента полимеразы. В результате создаются две двойные цепи вместо одной.
Так происходит процесс самокопирования ДНК: на ее позитивной цепочке синтезируется негативная, а на негативной — позитивная дополнительные цепочки. Две новые молекулы ДНК тождественны исходной и имеют такую же последовательность ступенек молекулярной лестницы.
Способность к самокопированию — свойство только нуклеиновых кислот, которым не обладают даже белки.
С помощью самокопирования осуществляется передача наследственных признаков, «записанных» на двойных спиралях ДНК. В одной молекуле нуклеиновой кислоты записано генетическое строение значительного количества различных белков. Это означает, что последовательность аминокислот в белке закодирована последовательностью нуклеотидов соответствующего участка молекулы нуклеиновой кислоты. Поскольку сведения о строении белков — самое главное, что должно быть передано потомкам при размножении, мы говорим о кодировании, то есть зашифровке наследственной информации — о биохимическом коде наследственности.
Разнообразие фенотипов, наблюдаемое у животных, связано с различием в их ДНК. Когда дефект приписывается воздействию единичного гена, значит, произошло изменение на биохимическом уровне в единственной молекуле белка. Скорее всего, при этом обнаружится и специфическое изменение в основной цепочке ДНК. Так, из-за мутации вместо порядка AAA (три молекулы аденина) может возникнуть GAA, то есть на месте одной из молекул аденина появится молекула гуанина, что приведет к изменению кода, а значит, и наследственной информации. Чтобы доминантные гены вызвали биохимическую реакцию для фенотипического проявления признака, достаточно правильного сообщения, хранящегося только в одной из хромосом — только отцовской или материнской, тогда как для проявления рецессивного признака оно должно содержаться в обеих хромосомах — как в отцовской, так и в материнской.
Подробно об этом написали в 1966 году Д.Ф. Паттерсон и У. Медуэй [710]. В своем труде за 1975 год [707] Д.Ф. Паттерсон приводит в качестве примера серповидноклеточную анемию (наследственная гемолитическая анемия) у человека, показав, что сложная клиническая картина этого тяжелого заболевания (периодический гемолиз, анемия, изменения со стороны костей и т. д.) связана с тем, что на определенной стадии биохимической реакции место валина занимает глутаминовая кислота, что, в свою очередь, приводит к замене аденина на определенном участке т-РНК тимином.
Поскольку конечный продукт серии биохимических реакций зависит от правильного течения каждой из них, в самих сериях могут произойти изменения на разных этапах, способные привести к тому же самому результату. Так, независимо от причин, приводящих к гемофилии, заболевание проявляется в нарушении образования фактора VIII, ускоряющего свертывание крови. Ген, детерминирующий синтез этого фактора, находится в участке Х-хромосомы, не имеющей гомолога, и представлен двумя аллелями — доминантным нормальным и рецессивным мутантным. Нарушение любой из различных реакций в биохимической цепочке приведет к гемофилии, но на каждом этапе за них отвечают разные гены — одни из них сцеплены с полом, другие же — аутосомные.
Наблюдая циклическую нейтропению (резкое уменьшение числа нейтрофилов в периферической крови с преобладанием юных форм) у длинношерстной колли, мы обнаруживаем у нее биохимические нарушения, не позволяющие нейтрофилам стать зрелыми, а также плейотропный эффект ослабления окраса шерсти, так называемый синдром серой колли, также связанный с изменением биохимических реакций, воздействующих на лейкоциты и формирование пигмента в волосах.
Биохимическая генетика — самостоятельный раздел науки, хотя заводчиков, разумеется, больше интересуют не биохимические реакции, происходящие в организме животного, а более простые механизмы наследования. Если мы знаем, что конкретный дефект связан с дупликацией рецессивной аллели, то можем спланировать программу племенного разведения, и лежащие в основе всего этого биохимические процессы нас вовсе не интересуют. Как бы мы ни представили себе гены (в виде ДНК или нитки бисера), математика генетических коэффициентов от этого не изменится.
|
| | | | Гость
Гость
| | Тема: Re: Генетика собак 17/1/2014, 1:45 am | |
| Некоторые признаки, называемые зависимыми от пола, определяются генами, находящимися не в Х-хромосомах, а в одной из пар аутосом. Однако проявление таких признаков зависит от пола животного (независимо от сцепления с полом), что обычно связано с действием мужских или женских половых гормонов (в рецессивном состоянии признак отсутствует независимо от пола).
Типичными примерами являются все вторичные половые признаки — комолость у некоторых пород овец, раннее облысение у людей, яйценоскость кур и наследование окраса у эр- ширских молочных коров, как показано в таблице 4.
ТАБЛИЦА 4
Зависимость окраса эрширских молочных коров от пола
Генотип Фенотип
Быки Коровы
ММ Махагон Махагон
Mm Махагон Красный
mm Красный Красный
Некоторые признаки, ограниченные полом, проявляются только у самок, как, например, молочность, или у самцов, как крипторхизм, хотя носителями их могут быть и те и другие (так как они полигенные, а контролирующие их гены расположены в любых парах аутосом). Это осложняет генетические исследования и в некоторых случаях означает, что при селекции следует руководствоваться качеством потомства в целом, а не отдельно взятого животного, так как по одному полу невозможно сделать правильные выводы.
Некоторые признаки, зависящие от пола, проявляются и у самцов, и у самок, но у одних из них они встречаются чаще, а у других — реже. Так, заячья губа и подагра, встречающиеся у людей, чаще всего наблюдаются все-таки у мужчин, а вот spina bifida (расщелина позвоночника — врожденная аномалия, характеризующаяся неполным закрытием позвоночного канала из-за отсутствия дуги нескольких позвонков, главным образом в поясничной области) и дисплазия тазобедренного сустава — у женщин.
Наследование ограниченных полом признаков осуществляется в соответствии с законами Менделя.
НЕПОЛНАЯ ПЕНЕТРАНТНОСТЬ
При обсуждении доминантных и рецессивных аллелей предполагалось, что конкретный признак проявляется у животного в присутствии одного-единственного доминантного аллеля. Рецессивный же аллель для его проявления должен быть в гомозиготном состоянии. В этом случае мы имеем дело с так называемой полной (абсолютной) пенетрантностью {от лат. penetrare — проникновение). В некоторых же случаях, чаще всего у гетерозиготных особей (Аа), доминантный ген А в 75% случаев может вести себя как доминантный, а в 25% случаев — как рецессивный. В этом случае речь идет о неполной, а 75%-ной пенетран- тности. Причиной тому чаще всего служит полигенное наследование признака.
При неполной пенетрантности заводчик сталкивается с определенными трудностями, ибо на первый взгляд результаты вязок не соответствуют законам Менделя, что связано с недостаточным количеством потомства для его анализа.
ФЕНОКОПИИ
Заводчики часто ошибочно полагают, что наследственность зависит только от генотипа, забывая о факторах индивидуального развития организма и влияния внешней среды. Так, травма суки во время щенности может значительно повлиять на развитие эмбриона, но не на его генотип, в отличие от внешних воздействий, способных вызвать мутации у плода. Особь с измененным генотипом называется фенокопией.
Пожалуй, самым трагичным примером фенокопий у человека можно назвать случаи, связанные с применением беременными женщинами в 60-х годах талидомида — транквилизатора с тератогенным действием, приводящего к фокомелии или ахе- роподии у новорожденных (укорочению или отсутствию конечностей).
Примером фенокопии служат и мягкие хрящи у собак со стоячими в норме ушами. Конечно, их можно поставить за счет фиксации ушных раковин, но такие собаки стойко будут передавать полустоячие уши своему потомству.
ВАРИАЦИИ ЧИСЛА ПОЛОВЫХ ХРОМОСОМ
Я уже писал, что в норме кариотип собак включает 78 хромосом, образующих 39 пар, включая одну половую: XX — у сук и XY — у кобелей. При изучении генного баланса, генетики обнаружили, что, даже если он и нормальный, в нем все равно встречаются отклонения. Вариации числа половых хромосом могут быть связаны с целым набором хромосом (эуплоидия) или с одной хромосомой (анэуплоидия).
Пожалуй, самым известным примером такого явления служит синдром Дауна у человека, когда в двадцать первой паре хромосом (в норме их 23) появляется лишняя хромосома. Это происходит на определенной стадии формирования гамет и чаще наблюдается среди старородящих, как правило сорокалетних матерей.
Возможно, подобные хромосомные аберрации происходят чаще, чем нам кажется, но мы не так часто видим их проявле-
Рис. 12. Типы нехваток хромосом
1 — концевая (терминальная, или дефишиенси); 2 — две концевые нехватки, приводящие к образованию кольца; 3 — внутрихромосомная нехватка (делеция)
ние из-за гибели гамет или ранней смертности плода. Болезнь Дауна (одна из форм олигофрении) — один из тех случаев, когда аберрация не настолько серьезная с генетической точки зрения, чтобы вызвать раннюю смерть эмбриона. А так как подобные аномалии встречаются в собаководстве чрезвычайно редко, мало кто из ученых придает им большое значение.
Второй тип — дупликация (удвоение) хромосом. В этом случае одновременно с возникновением нехватки в результате разрыва хромосомы и потери участка внутри одной хромосомы могут накапливаться идентичные участки, несущие одни и те же гены. При дупликации происходит фенотипическое изменение признаков.
Если в норме каждый участок хромосомы представлен одной дозой гена, то при удвоении или утроении несущего эти гены участка доза гена увеличивается в два или три раза. То есть у сук имеется не диплоидный набор (2Х:2А, то есть половой индекс, или соотношение между Х-хромосомами и аутосомами, равен единице), а три половые хромосомы — XXX, из-за чего они называются сверхсуками (они могут быть диплоидными — ЗХ:2А с
половым индексом 1,5 или триплоидными — ЗХ:ЗА). У таких сук наблюдается нарушение астральных циклов (как правило, они стерильные), а если и нет, то при их вязке с нормальными кобелями рождаются интерсексы (2Х:ЗА с половым индексом 0,66) мужского или женского типов — гермафродиты. У кобелей комплекс XXY (синдром Клайфельтера) также сопровождается явлениями инфантилизма из-за недоразвития семенников и азооспермии. Лишняя Х-хромосома у котов обусловливает появление в помете самок черепахового окраса, реже самцов с половым набором XXY, страдающих стерилитетом.
Животные с лишней половой хромосомой, как правило, стерильные. Типичный пример — мул (гибрид кобылы, имеющей 64 хромосомы, и осла, у которого их 62), кариотип которого включает 63 хромосомы. Реже у сук имеется всего лишь одна половая хромосома — Х0, что проявляется в синдроме Шерешевского-Тёрнера (с явлениями инфантилизма и отсутствием яичников). Кобели с набором XY:3A (половой индекс 0,33), вместо XY:2A (половой индекс равен 0,5), чрезмерно мускулистые, грубого сложения, с массивным костяком и называются сверхкобелями. Они также бесплодны.
Иногда у кобелей бывает лишняя Y-хромосома, в этом случае у них, как и у мужчин с набором XYY, отмечается неоправданная и неуправляемая агрессия.
НЕКОТОРЫЕ БИОХИМИЧЕСКИЕ АСПЕКТЫ
В 1977 году М. Бёрнс [141], обсуждая мою книгу «Немецкая овчарка», опубликованную в 1976 году, высказала мнение, что я придерживаюсь несколько старомодного подхода к основам генетики. Это критическое замечание остается справедливым и по отношению к данной книге. С моей точки зрения, заводчики лучше усваивают генетику, объясняемую с точки зрения доминантных и рецессивных признаков, чем в свете более современных биохимических процессов. Однако замечание М. Бёрнс вполне справедливо, и для упрощенного объяснения действия генов я все-таки коротко поясню некоторые биохимические аспекты.
До этого я привел вам в качестве аналогии нитку бисера для иллюстрации понятия генов и хромосом. В строгом смысле она некорректна, ибо гены — не изолированные объекты, а составляющие части хромосомы. Вся информация, необходимая для определения наблюдаемых нами в отдельной особи
Один нуклеотид-
Один нуклеотид
Сахаро фосфат- Пары комплементарных " (Сахарофосфатный остов t оснований ный остов
Одна полинуклео- н-связь Одна полинуклео- тидная цепь . . тидная цепь
Рис. 14. ДНК (схематическое изображение развернутых цепей) А — аденин I
I Сахарофосфатный '.ОСТОВ
Две антипараллельные цепи
2 нм
Рис. 13. Схематическое изображение структуры ДНК. На один полный оборот спирали приходится 10 пар оснований
3,4 км
Основания-
кольца
Г — гуанин I Т — тимин ] Ц — цитозин I
одно кольцо
О фосфа°тДНЫе СВЯЗИ \ / Дезоксирибоза (сахар)
признаков, содержится в структуре ДНК, из которой состоят гены. Сама ДНК — полимер, менее сложный по сравнению с белками, состоящий из сахара (дезоксирибозы), остатков молекул фосфорной кислоты и четырех азотистых оснований (пу- риновых — аденина, гуанина и пиримидиновых — цитозина и тимина). Молекула нуклеиновой кислоты представляет собой цепочку мономеров, специфических для этих кислот — нукле- отидов. Строение определенной молекулы нуклеиновой кислоты характеризуется определенной последовательностью нукле- отидов. Каждый нуклеотид состоит из трех разных элементов: азотистого основания, углеводного компонента и фосфорной кислоты.
Для нуклеотидов ДНК характерна способность образовывать самые длинные молекулы из всех известных. Вероятно, каждый ген в хромосоме представляет собой участок ДНК, содержащий сотни или тысячи нуклеотидов.
В 1953 году американский биохимик Джеймс Дьюи Уотсон и английский биофизик и генетик Фрэнсис Харри Комптон Крик выдвинули гипотезу, что молекула ДНК представляет собой двойную спираль, напоминающую лестницу со ступеньками из азотистых оснований, соединенных строго специфически (аденин с тимином, а гуанин с цитозином), перилами которой служат остатки фосфорной кислоты и пентозного сахара. Они же создали модель молекулы ДНК, а в 1962 году они вместе с английским биофизиком Морисом Уилкинсом впервые получили высококачественные рентгенограммы молекулы ДНК (см. рис. 13). Цепочки полинуклеотидов этой спирально завитой двойной молекулы расположены так, что сахар и фосфорные группы лежат снаружи молекул, а основания, соединенные между собой водородными связями, — внутри молекул (см. рис. 14).
В отдельной цепочке из двадцати аминокислот хранится информация, отвечающая за синтез определенного белка, считываемая информационной, или матричной, РНК и переносимая транспортной РНК к рибосомной РНК. В определенных условиях двойная цепь ДНК разрывается на месте водородных связей и возле каждой цепи строится новая комплементарная цепь с помощью фермента полимеразы. В результате создаются две двойные цепи вместо одной.
Так происходит процесс самокопирования ДНК: на ее позитивной цепочке синтезируется негативная, а на негативной — позитивная дополнительные цепочки. Две новые молекулы ДНК тождественны исходной и имеют такую же последовательность ступенек молекулярной лестницы.
Способность к самокопированию — свойство только нуклеиновых кислот, которым не обладают даже белки.
С помощью самокопирования осуществляется передача наследственных признаков, «записанных» на двойных спиралях ДНК. В одной молекуле нуклеиновой кислоты записано генетическое строение значительного количества различных белков. Это означает, что последовательность аминокислот в белке закодирована последовательностью нуклеотидов соответствующего участка молекулы нуклеиновой кислоты. Поскольку сведения о строении белков — самое главное, что должно быть передано потомкам при размножении, мы говорим о кодировании, то есть зашифровке наследственной информации — о биохимическом коде наследственности.
Разнообразие фенотипов, наблюдаемое у животных, связано с различием в их ДНК. Когда дефект приписывается воздействию единичного гена, значит, произошло изменение на биохимическом уровне в единственной молекуле белка. Скорее всего, при этом обнаружится и специфическое изменение в основной цепочке ДНК. Так, из-за мутации вместо порядка AAA (три молекулы аденина) может возникнуть GAA, то есть на месте одной из молекул аденина появится молекула гуанина, что приведет к изменению кода, а значит, и наследственной информации. Чтобы доминантные гены вызвали биохимическую реакцию для фенотипического проявления признака, достаточно правильного сообщения, хранящегося только в одной из хромосом — только отцовской или материнской, тогда как для проявления рецессивного признака оно должно содержаться в обеих хромосомах — как в отцовской, так и в материнской.
Подробно об этом написали в 1966 году Д.Ф. Паттерсон и У. Медуэй [710]. В своем труде за 1975 год [707] Д.Ф. Паттерсон приводит в качестве примера серповидноклеточную анемию (наследственная гемолитическая анемия) у человека, показав, что сложная клиническая картина этого тяжелого заболевания (периодический гемолиз, анемия, изменения со стороны костей и т. д.) связана с тем, что на определенной стадии биохимической реакции место валина занимает глутаминовая кислота, что, в свою очередь, приводит к замене аденина на определенном участке т-РНК тимином.
Поскольку конечный продукт серии биохимических реакций зависит от правильного течения каждой из них, в самих сериях могут произойти изменения на разных этапах, способные привести к тому же самому результату. Так, независимо от причин, приводящих к гемофилии, заболевание проявляется в нарушении образования фактора VIII, ускоряющего свертывание крови. Ген, детерминирующий синтез этого фактора, находится в участке Х-хромосомы, не имеющей гомолога, и представлен двумя аллелями — доминантным нормальным и рецессивным мутантным. Нарушение любой из различных реакций в биохимической цепочке приведет к гемофилии, но на каждом этапе за них отвечают разные гены — одни из них сцеплены с полом, другие же — аутосомные.
Наблюдая циклическую нейтропению (резкое уменьшение числа нейтрофилов в периферической крови с преобладанием юных форм) у длинношерстной колли, мы обнаруживаем у нее биохимические нарушения, не позволяющие нейтрофилам стать зрелыми, а также плейотропный эффект ослабления окраса шерсти, так называемый синдром серой колли, также связанный с изменением биохимических реакций, воздействующих на лейкоциты и формирование пигмента в волосах.
Биохимическая генетика — самостоятельный раздел науки, хотя заводчиков, разумеется, больше интересуют не биохимические реакции, происходящие в организме животного, а более простые механизмы наследования. Если мы знаем, что конкретный дефект связан с дупликацией рецессивной аллели, то можем спланировать программу племенного разведения, и лежащие в основе всего этого биохимические процессы нас вовсе не интересуют. Как бы мы ни представили себе гены (в виде ДНК или нитки бисера), математика генетических коэффициентов от этого не изменится.
Вполне понятно, что, если произвольно выбрать пять сук, измерить их высоту в холке и счесть результат типичным для породы, шанс на ошибку будет высоким. Вернувшись к гистограмме 3, мы получим ошибку, равную 0,09 см, то есть разброс по средней высоте в холке в пределах 60,01—60,19 см. В настоящее время средняя высота немецких овчарок в холке составляет в Германии 58 см, а девиация крайне мала. Это говорит о том, что мы должны рассматривать популяцию как постоянно изменяющуюся, так что нам следует постоянно анализировать изменения, происходящие в ней, строя новые и новые вариационные кривые. Мы не знаем точно, с каким числом генов имеем дело, зато можем математически оценить каждый признак и определить степень его влияния на изменчивость, что крайне важно в разведении.
|
| | | | Гость
Гость
| | Тема: Re: Генетика собак 17/1/2014, 1:45 am | |
| Итак, мы рассмотрели понятие изменчивости или разнообразия признаков. Все полигенно контролируемые признаки выстраиваются в вариационный ряд, включая высоту в холке, массу тела взрослой особи, угол наклона лопатки и стати экстерьера, оцениваемые на ринге. Но то, что мы видим и измеряем, — фенотипическая изменчивость, которая зависит не только от генотипической (наследственной) изменчивости, но и от воздействия на организм факторов внешней среды.
Фенотипическая изменчивость не затрагивает генотипа и не сохраняется при половом размножении. Она выражается в разнообразии особей с единым генотипом. Один из видов этой изменчивости — онтогенетическая, которая наблюдается в процессе индивидуального развития с закономерными изменениями морфологических, физиологических, биохимических и других особенностей организма. Другой вид — модификационная изменчивость, наблюдающаяся среди особей с единым генотипом, но живущих в разных условиях.
Генотипическая изменчивость — изменение признаков организма, обусловленное изменением генотипа и сохраняющееся в ряду поколений. Она может возникать путем скачкообразных изменений признаков (мутационная изменчивость) или путем новых комбинаций уже существующих признаков в результате скрещивания (комбинативная изменчивость). Генотипическая изменчивость условно делится на два компонента — аддитивный и неаддитивный, который в свою очередь состоит из доминантного и эпистатического.
Для вычисления квадратического отклонения в фенотипе обозначим вариабельные компоненты изменяемого признака следующим образом: фенотипический — Р, генотипический — G, экологический — Е, аддитивный — А, доминантную девиацию — D, а эпистатическую — I. В этом случае мы получим следующую формулу:
о2 = о2 + а2 = о2 + о2 + а2 + о2
Up uG>uE и д 1 и D 1 и , 1 и
Относительное значение различных компонентов будет зависеть от рассматриваемого конкретного признака и популяции, в которой он изучается. Когда речь идет о простых признаках, экологический компонент может оказаться настолько незначительным, что его можно проигнорировать, но более сложные признаки всегда подвергаются воздействию окружающей среды, да и генетический эффект может быть разнообразным за счет аддитивного или неаддитивного взаимодействия генов.
Аддитивный компонент — часть изменчивости, наиболее интересная для заводчика, так как является основной причиной фенотипического сходства между родственниками, поэтому он и называется племенной ценностью. Неаддитивный компонент O2d + о2,) сложен в рассмотрении, но его действие основано на взаимодействии с геном в том или ином локусе (O2D) ИЛИ С рядом генов, расположенных в разных локусах (о2,). Иными словами, если проявление признака связано с взаимодействием специфических комбинаций генов в одном или нескольких локусах, неаддитивный компонент изменчивости будет довольно-таки высоким. Это означает, что признак, находящийся под контролем преимущественно неаддитивных генов, будет плохо поддаваться селекции. Объясняется это тем, что родители передают своему потомству набор генов, а не их специфические комбинации. Следовательно, животное, обладающее превосходным, но практически ненаследуемым признаком, не сможет воспроизвести его в потомстве.
Степень генетического контроля над признаком определя-
ется простым соотношением — —г, но ее значение не поможет
заводчику при низком значении компонента а2А. Это подводит нас к наследуемости признака, или Л2, указывающей, какая часть изменчивости находится под действием аддитивных генов, что предоставляет заводчикам потенциальную возможность добиться прогресса при селекции. Наследуемость признака определяется по формуле:
о . h2=-f. стР
Для заводчика, не занимающегося статистическим учетом и анализом вариационных рядов изменчивости, это может показаться довольно-таки сложным. При заводском или лабораторном разведении (при поголовье более ста животных) ведется обширная научно-исследовательская работа по выявлению наследуемости множества ценных признаков, на основании чего разрабатываются программы племенного разведения.
Таким путем было вычислено количественное значение наследуемости многих признаков, важных для разведения домашнего скота и лабораторных животных, в частности крыс и мышей.
Из всего вышесказанного можно заключить, что признаки, связанные с «пригодностью», имеют низкую наследуемость и ими можно пренебречь. «Пригодные» признаки включают большинство особенностей репродукции, например сроки оплодотворения, многоплодие или малоплодие, жизнестойкость и т. п. Их низкая наследуемость не означает, что они не находятся под генетическим контролем (это подтверждается тем, что для разных пород животных характерны свои сроки оплодотворения), а говорит о незначительном влиянии аддитивных генов, в результате чего трудно оценить племенную ценность животного. Низкая наследуемость указывает, что селекция по тому или иному признаку вряд ли даст какой-либо заметный результат. Селекцию, направленную на многоплодие, пришлось бы скрупулезно вести на протяжении многих лет, прежде чем средняя численность помета в породе увеличилась бы на одного щенка.
С другой стороны, признаки, связанные с ростом и развитием, скорее всего, обладают средним или высоким уровнем наследуемости. Шведские ученые провели исследование в Шведском центре армейских собак по наследуемости веса у двухмесячных щенков немецкой овчарки: у 45% кобелей и 34% сук, доказав, что, если бы заводчики решили заняться селекцией собак такого же возраста по прибавке в весе, она была бы и эффективной, и быстрой. То же самое можно сказать и о снижении веса в этом возрасте. И это притом, что прибавка веса в одном возрасте напрямую связана с его увеличением в другом. В 1928 году в Германии кобели немецкой овчарки имели высоту в холке около 64,4 см, а средний вес — 33,5 кг. К 1973 году они имели рост около 63,5 см, а вес — 37,2 кг, что свидетельствует о небольшом снижении средней высоты в холке и существенном (около 11%) увеличении веса за сорок пять лет. Отчасти это связано с совершенствованием методов кормления, а также с целенаправленной селекцией по типу, значительно отличающемуся от того, что преобладал в 20-х годах. Это стало возможным благодаря тому, что такие признаки, как высота в холке и вес, находятся под достаточно высоким контролем аддитивных генов.
Хотя оценить породный тип не так-то и просто, разумно предположить, что его наследуемость колеблется от умеренной до высокой, что позволило нам вывести большое многообразие пород. Циник может усомниться в разумности изменения экстерьера ряда пород, но никто не опровергнет ее эффективности.
Ученые изучили также генетику поведения собак, причем полученные ими данные свидетельствуют скорее о низкой наследуемости поведенческих реакций.
Концепция наследуемости может показаться довольно-таки сложной, но она, бесспорно, подчеркивает взаимосвязь родителей с их потомками, а также между сибсами (брата с сестрой) и полусибсами (полубрата с полусестрой), на которых базируются методы оценки изменений в потомстве по сравнению с родителями или корреляции между сибсами и полусибсами. Поскольку в разведении чаще используются кобели, большинство наследуемых признаков оценивается либо по их изменению у потомков по отношению к отцу, либо из корреляции между полусибсами по отцовской линии. Пронализировав признаки и построив график изменчивости признака, мы получим линию, отображающую, что потомки унаследовали лишь половину признаков отца, а на корреляционной — четверть.
|
| | | | Гость
Гость
| | Тема: Re: Генетика собак 17/1/2014, 1:46 am | |
| Из формулы о2р = о2с + О2е = о2А + O2d + о2, + О2е следует, что полная изменчивость конкретного признака, который мы можем видеть и измерить, зависит главным образом от двух факторов: от степени генетического контроля над ним и от воздействия на него окружающей среды.
Есть поговорка: «Происхождение наполовину зависит от того, что мы едим и чем дышим», и это весьма логично в том смысле, что реализация генетического потенциала собаки во многом зависит от среды ее обитания.
Большее или меньшее влияние факторов внешней среды на организм начинается с момента оплодотворения. Так, белый щенок может не перецвести из-за того, что во время щен- ности его мать получала низкокачественный рацион, или потому, что у него развился рахит. С другой стороны, оба эти фактора могут повлиять на другие гены, что не позволит ему стать хорошо развитым, крепкокостным кобелем с высоким выставочным потенциалом.
Большинство признаков, связанных с полигенией, испытывают более или менее сильное влияние внешней среды, включая не только питание во время внутриутробного, неонатального и ювенильного развития, результаты которого очевидны для всех опытных заводчиков, но и другие, менее явные факторы. Клинические и субклинические заболевания могут отрицательно сказываться на внешнем виде, породном типе и выживаемости собаки, а ранняя социализация положительно сказывается на ее дальнейшем поведении. Следующие аспекты — климат и температура окружающей среды. У собак, попавших в жаркие климатические условия, могут проявиться признаки, которых не бывает в умеренном климате, например кожные заболевания или увеличение мошонки.
Но кроме внешних, на собаку влияют и внутренние факторы, также способные вызвать у нее те или иные изменения. Наиболее наглядный из них — возраст, так как проявление многих признаков зависит именно от него, а именно: перецветание черной шерсти в голубую, происходящее у керри-блю-терьеров к возрасту года и восьми месяцев, постепенное укрепление ушного хряща и подъем стоячих ушей к четырехмесячному возрасту, постепенное потемнение радужной оболочки глаза, начиная с трехнедельного возраста, и такие нежелательные, как, например, HD, проявляющаяся по мере созревания собаки. Одни из этих признаков, например длина шерсти, проявляются довольно-таки рано, а такие, как PRA или врожденная эпилепсия, — к старости.
Половой диморфизм также зависит от генотипа, влияния внешних и внутренних факторов, в частности гормонального фона животного. Среди заболеваний также есть такие, которые чаще встречаются у какого-то одного пола.
И последняя причина изменчивости, которую можно причислить к факторам внешней среды, — ошибка в ее определении или расчете. Любая ошибка, допущенная нами при измерении высоты собак в холке, не отражается на самих животных, но влияет на наши расчеты в популяции. Подобные ошибки увеличивают математическое значение компонента общей изменчивости сг2Е, что приводит к уменьшению значения о2д. Таким образом, ошибки в оценках приводят к снижению значений наследуемости. Это относится не только к таким физическим параметрам, как рост и вес, но и к ошибкам, возникающим при неправильной классификации HD по степени ее тяжести.
Понятно, следует приветствовать такие расчеты, при которых ошибка приближается к нулю. Если породный клуб желает внести в стандарт физические параметры, ему следует исключить признаки, связанные с возрастом, и использовать большую выборку собак одного возраста по линейным признакам, чтобы вывести среднее значение каждого из них.
В кинологических кругах никогда не поощрялись математические расчеты, и, безусловно, сам по себе статистический анализ линейных признаков собственного поголовья еще никого не сделал хорошим заводчиком. Но пока все признаки, которые мы хотим закрепить или свести на нет благодаря селекции, не будут документально зафиксированы, не стоит уповать на то, что нам удастся оценить, насколько правильный путь нам удалось для этого выбрать.
КОРРЕЛЯЦИЯ
Корреляция — зависимость разнообразия одного признака от изменчивости другого. Биометрия позволяет установить наличие или отсутствие связи между отдельными признаками, а также ее направление и силу. Отсутствие корреляции показано на графике 1. Корреляция по направлению может быть положительной (с увеличением одного признака связанный с
г = 0.03
Признак А
J 1 I I I
Признак С ~~"""
Признак В
г = 0.99
Признак А
t
г =-0.79
Признак С
J 1 I ' I
Признак D
График 1. Отсутствие корреляции
Точки — отдельные животные с ценными признаками А и С Прямая линия — наилучшее сочетание двух признаков
График 2. Положительная корреляция
Точки — отдельные животные с ценными признаками А и В Прямая линия — наилучшее сочетание двух признаков
График 3. Негативная корреляция
Точки — отдельные животные с ценными признаками С и D Прямая линия — наилучшее сочетание двух признаков
ним другой также увеличивается — график 2) и отрицательной или разнонаправленной (с увеличением одного признака другой уменьшается — график 3). Показателем связи между признаками служит коэффициент корреляции (с), абсолютная величина которого показывает силу связи, а знак перед ним — ее направление. При отсутствии связи с = 0, если она слабая, с < 0,3, при средней с = 0,3—0,7, а при сильной с > 0,7.
Заводчики ведут отбор не по одному, а по целому ряду признаков. Для того чтобы свести селекцию к максимуму, следует познакомиться с соответствующими корреляциями, поскольку бесполезно вести отбор по какому-то одному незначительному признаку, если это приведет к регрессу более важного и необходимого для породы.
Хотя нам мало что известно о корреляциях между признаками, логично высказать несколько предположений. Если мы, например, увеличим путем селекции высоту у собак в холке, то вслед за этим обязательно повысим их вес, так как между размером скелета и весом существует положительная корреляция. Но в ходе подобной селекции могут появиться и другие изменения, например многоплодие, а также спрямление углов задних конечностей и загруженности крупа. Если мы проведем обратную селекцию и попробуем получить внутрипородную миниатюрную или той-разновидность, она будет менее эффективной, так как мы столкнемся с дистоцией (тяжелыми родами) и малоплодием.
Причина корреляции может быть двойственной. С одной стороны, это полимерия, когда гены, отвечающие за проявление одного и того же признака, находятся в одной хромосоме, то есть в группе сцепления. Чем меньше расстояние между их локусами и чем больше в хромосоме однозначных доминантных генов, тем больше вероятность и выраженность их совместного (кумулятивного) действия. С другой стороны, это может быть связано с явлением плейотропии, когда один ген влияет на несколько признаков. Плейотропия является одной из причин генетической корреляции. Менделевское расщепление в F2 при дигибридном скрещивании 9:3:3:1 происходит лишь при соблюдении ряда условий, прежде всего при независимом наследовании признаков и отсутствии взаимодействия между генами. Из-за того, что большинство признаков определяются взаимодействием аллельных генов (неполное доминирование и кодо- минантность) и неаллельных генов (комплементарность, эпистаз и полимерия), в F2 могут наблюдаться необычные отношения — 9:3:4; 9:7; 13:3; 12:3:1 и 15:1. Наверняка именно этим и объясняется взаимосвязь между размером скелета и массой тела, которые контролируются несколькими генами, некоторые из которых влияют на оба эти признака в одном и том же направлении.
Плейотропия имеет важное, но не ведущее значение в селекции. Так, некоторые заводчики доберманов заявляли, что лучшие собаки американских линий — красно-подпалые. Но наследование окраса у доберманов подчиняется простым законам Менделя (см. табл. 3), так что неразумно предполагать, что серия простых генов тесно связана с множеством других, отвечающих за появление высококлассного супертипа. Подобные заявления возникают из-за пренебрежительного отношения к документации. Если бы она велась и охватывала анализ наследственности и изменчивости признаков у достаточно большого (статистически достоверного) числа доберманов, они, скорее всего, были бы опровергнуты. Когда складывается впечатление об истинной взаимосвязи окраса с каким-то важным породным признаком, как правило, выясняется, что он был получен от одного из близстоящих выдающихся предков, большинство потомков которого унаследовали его расцветку вместе с множеством физических достоинств. Если не продолжать селекцию по окрасу и типу, то взаимосвязь между ними постепенно исчезнет.
|
| | | | Гость
Гость
| | Тема: Re: Генетика собак 17/1/2014, 1:46 am | |
| Многие признаки животного можно оценить лишь раз. Например, вес новорожденного измеряется сразу после его рождения. Можно взвесить всех новорожденных в помете и вычислить их средний вес, но и при этом вы все равно прибегнете лишь к однократному измерению признака. И наоборот, определение скорости, с какой грейхаунд бежит по беговой дорожке, можно измерять множество раз, чтобы составить более точное представление о его резвости.
Аналогичным образом можно выявить среднее число щенков в помете и многоплодных в популяции сук после их щенения, чтобы выбрать для племенной работы наиболее плодовитых из них. Учитывая, что каждая щенная сука приносит разное количество щенков, статистический анализ позволяет определить частоту повторяемости признака (г) в популяции, то есть корреляцию между его измерениями, в данном случае — численности помета. Частота появления у суки одинаковых по численности пометов зависит от ее генотипа и воздействия на гены, отвечающие за многоплодие, от внешних и внутренних факторов.
До сих пор мы рассматривали разнообразные факторы внешней среды в целом, но когда речь идет о повторяемости признака, их стоит разделить на два компонента: o2Eg — общие для популяции, то есть ведущие к одним и тем же изменениям в численности их пометов, и G2Es — специфические, касающиеся конкретной суки и ее плодовитости.
Таким образом, частота повторяемости признака отражает взаимосвязь специфических воздействий разнообразных факторов внешней среды (o2£s) с частными и общими генетическими изменениями и влиянием общих факторов внешней среды (O2G + O2EG), что выражается в следующей формуле:
Следовательно, частота повторяемости признака отражает соотношение изменчивости, возникшей за счет перманентных или нелокализованных различий между индивидуумами. Она будет меняться в зависимости от признаков и породы, но ее следует учитывать в селекции, так как:
1) частота повторяемости признака устанавливает максимальное ограничение, или верхний предел наследуемости, как в узком
°2а O2g
смысле этого слова ), так и в широком ее легче оценивать, чем два этих отношения, и можно вычислить и без них;
2) зная частоту повторяемости признака, мы можем выяснить, насколько часто его следует измерять.
Допустим, мы оцениваем численность помета, зная среднюю плодовитость сук данной породы. Сведя значение специфических
влияний различных факторов внешней среды к отношению ^ , где п — число пометов от каждой суки, мы получим следующую формулу:
1
°2Р(„) = °2с + °2Eg +й°2Ее.
Нам крайне выгодно свести влияние внешней среды на изменение признака на нет. Высокая частота повторяемости признака означает, что на него практически не влияют специфические факторы внешней среды, а значит, и нет никакой необходимости в его многократных измерениях. И наоборот, низкая частота повторяемости признака означает, что для успешного его закрепления нам требуются многократные измерения. Заметим, что частота повторения признака может быть высокой при низкой аддитивной изменчивости, а значит, и низкой наследуемости. Вполне вероятно, что это касается и численности щенков в помете. То есть сама сука может быть много- или малоплодной, что вовсе не означает, что она передаст эти качества своему потомству.
Данные об оценке частоты повторяемости признаков в трех пометах от каждой из 70 сук бернских зенненхундов, приведенные в табл. 5, были взяты мной из работы немецкого ученого Г. Кайзера [473]. Количество щенков в помете было 1 — 15, в трех пометах — 5—41, в первом помете в среднем их оказалось 7,36, во втором — 7,47, а в третьем — 6,8. Частота повторяемости признака равнялась 0,71, то есть была достаточно высокой, причем ее можно отнести и к другим породам собак с подобным средним количеством щенков в пометах.
ТАБЛИЦА 5
Расчет частоты повторяемости признака на примере плодовитости сук бернского зенненхунда
Необработанные данные, приведенные Г. Кайзером в 1971 году
Количество Число Количество Количество
щенков пометов щенков в трех щенных
в помете пометах сук
1 5 5 1
2 7 8 1
3 13 11 2
4 10 12 2
5 18 14 3
6 29 15 2
7 34 16 6
8 31 17 1
9 16 18 4
10 18 19 3
11 16 20 7
12 7 21 3
13 3 22 4
14 2 23 4
15 1 24 4
25 4
26 5
27 3
28 2
29 2
30 1
31 1
32 1
33 2
37 1
41 1
Анализ изменчивости по М.Б. Уиллису
Всего пометов (N) — 210 Всего сук (D) — 70 Всего щенков (Р) — 1514
Источник изменчивости Составляющие Сумма квадратов Средняя квадратичная
Групповой (D) 69 ka> D + a2w 1008,11 14,61
Индивидуальный (W) 140 244,67 1,748
Общая изменчивость 209 1252, 78
Составная групповой изменчивости в трех пометах = (14,61 — 1,748):3 = 4,287.
г = 4,287:(4,287 + 1,748) = 0,71.
ПОРОГОВЫЕ ЗНАЧЕНИЯ ПРИЗНАКОВ
До сих пор мы рассматривали полигенные признаки с точки зрения их непрерывной изменчивости, но иногда мы сталкиваемся и с их дискретной изменчивостью. В двух случаях некоторые из полигенных признаков все же могут изменяться в ограниченных пределах. Так, жизнеспособность щенка может зависеть от сложного взаимодействия целой серии генов, экспрессия которых визуально проявится в том, родится ли он живым или мертвым.
На первый взгляд кажется, что подобные признаки подчиняются простым законам Менделя (как в случае с черным и печеночным окрасами), а не сложной полигенной наследственности, но из генетики мы знаем, что в большинстве случаев это именно так.
Ключ к пониманию подобных признаков дает концепция их порогового значения, когда они доходят до той крайней точки, в которой переходят от одного своего выражения к другому, например от жизни к смерти.
Гистограмма 6. Пороговое значение фенотипического проявления полидактилии у морских свинок в зависимости от количества генов, отвечающих за нее, в их генотипе по М.У. Стрикбергеру, 1968 г.
пороговое значение признака уровень
3 — трехпалые свинки; 4 — четырехпалые свинки; 5 — поголовье 1 (S,); 6 — поголовье 2 (S2) Р — родители N — трехпалые гибриды
Классическим примером подобного феномена служат результаты, полученные С. Райтом [1029] в 1934 году, при исследовании полидактилии, встречающейся и у собак, в двух поголовьях морских свинок, в одном из которых у животных было по три пальца на задних конечностях, а в другой — по четыре1. После скрещивания животных первого поголовья со вторым С. Райт получил трехпалых гибридов первого поколения (F,), после чего повязал их между собой, обнаружив, что в F2 родились и четырехпалые гибриды. Кроме того, он провел кросс трехпалых свинок из F, с четырехпалыми из второго поголовья, а трехпалых свинок из-под него вновь пустил в разведение со свинками второго поголовья (см. гистограмму 6). Проведя анализ, С. Райт понял, что полидактилия передается через рецессивный ген согласно простому закону Менделя.
Со временем С. Райту удалось выяснить, что за полидактилию отвечает восемь генов, причем фенотипически она проявляется при их накоплении в генотипе, начиная с пяти. Несмотря на то, что С. Райт изучал здоровых и четырехпалых свинок, их изменчивость по данному признаку соответствовала нормальной вариационной кривой (см. гистограмму 6). В генотипе свинок первого, фенотипически нормального, трехпалого поголовья либо не было вообще, либо имелось до двух генов полидактилии, а в генотипе второго, четырехпалого, их было 5—8. У гибридов F, в генотипе было всего 2—4 гена полидактилии, так что фенотипически они выглядели трехпалыми, и только достигнув порогового числа — 5, в помете появятся четырехпалые свинки.
Если мы имеем дело с контролируемыми признаками, как в этом примере, то единственный способ оценить их — получить различную степень их экспрессии в популяции. Согласно Д.С. Фалконеру [269], зная, что у 60% особей в популяции проявляется одно видоизменение признака, а у оставшихся 40% — другое, можно определить его пороговое значение.
' В норме у морской свинки по четыре пальца на передних и по три — на задних конечностях.
|
| | | | Злюсенький Admin
Admin

Сообщения : 11121
сказали СПАСИБО : 67
Возраст : 46
Откуда : Челябинск
| | Тема: Re: Генетика собак 17/1/2014, 11:06 am | |
| Офигееееееееть......... Позже почитаю)))))) | |
| | | | Muftocka.
Канаровед


ПРЕДУПРЕЖДЕНИЯ : "Не говори Богам, что ты в беде. Говори беде, что ты с Богами".
Сообщения : 4330
сказали СПАСИБО : 8
Возраст : 47
Откуда : Северная Европа. Юрмала
| | Тема: Re: Генетика собак 28/2/2016, 2:38 pm | |
| Основные понятия генетики о генетике  Племенное разведение собак Предметом изучения генетики являются два неразрывных свойства всех живых организмов — наследственность и изменчивость. Изменчивость представлена многообразием форм внутри каждого вида, породы и даже одного помета. Но в то же время все представители одного вида и одной породы имеют несомненное сходство обеспечиваемое наследственностью. Для каждого вида животных характерен набор хромосом определенной формы, составляющий кариотип. Половые клетки содержат половинный набор хромосом, называемый гаплоидным, который принято обозначать — n. Яйцеклетка образованная слиянием двух половых клеток содержит в два раза большее количество хромосом, так называемый, диплоидный набор — 2n. Каждая пара хромосом диплодного набора представлена, гомологичными хромосомами, одна из которых получена от отца, а другая от матери. Диплоидный набор собаки представлен 78 хромосомами. Все наследственные свойства и признаки определяются вполне определенными независимыми друг от друга материальными единицами — генами. Каждый ген занимает строго определенное место в строго определенной хромосоме, называемое локусом. В силу парности хромосом в клетках, гены хромосомном наборе также представлены парами. Гены, расположенные в одном локусе называются аллельными или аллелями. Гены могут претерпевать изменения в своем строении — мутировать, вследствие этого изменяются внешние проявления признака, за который отвечает данный аллель. Особи, получившие от отца и матери одинаковые аллели одного локуса, называются гомозиготными, а разные — гетерозиготными по данному признаку. Гены и аллели принято обозначать буквами латинского алфавита, например А, F, tfm и т. п. Взаимодействие аллельных генов Находясь в гетерозиготном состоянии аллели определенным образом взаимодействуют друг с другом. В случае, когда один них полностью подавляет действие другого, называется полным доминированием. Доминантный ген обычно обозначается заглавной буквой латинского алфавита. При полном доминировании гетерозиготные особи Aa имеют такой же внешний вид или фенотип, как и гомозиготные по доминантному аллелю АА. Это означает, что для проявления доминантного признака достаточно одного доминантного аллеля, что обозначается как A-. Если гетерозиготные особи отличаются по фенотипу от гомозиготных и имеют промежуточный фенотип, то говорят о неполном или промежуточном доминировании. Например, при скрещивании колли светло-соболиного окраса с колли трехцветного окраса получаются щенки темно-соболиного окраса. При сверхдоминировании — у гибридов первого поколения наблюдается гетерозис — явление превосходства потомства над родительскими формами по жизнеспособности, энергии роста, плодовитости. Так гибриды, полученные при скрещивании диких серых крыс — пасюков с белыми лабораторными, внешне похожи на пасюков, но значительно крупней и плодовитее чем последние. При кодоминировании у гибридной особи в равной мере проявляются оба родительских признака. По типу кодоминирования наследуется большинство антигенных факторов довольно многочисленных систем групп крови. В тех случаях, когда анализируется поведение признаков, обусловленных одной парой аллелей, например черного B и коричневого b окраса, говорят о моногибридном скрещивании. Скрещивание особей, отличающихся по двум парам признаков, называется дигибридным, по трем — тригибридным, по многим — полигибридным скрещиванием. Законы Менделя Еще в прошлом веке Грегором Менделем были показаны закономерности передачи признаков при скрещивании. Он сформулировал следующие законы: I закон Менделя — закон единообразия гибридов первого поколения. Скрещивание между собой особей, гомозиготных по разным аллелям одного локуса, приводит к рождению гетерозиготных потомков, одного фенотипа. Так при скрещивании между собой гомозиготных черных BB и коричневых bb собак все щенки получаются черными Bb. Особей, относящихся к родительскому поколению обозначают латинской буквой Р. Гибридов первого поколения — F1, гибридов второго поколения F2, гибридов третьего — F3 и т. д. II Закон Менделя — закон расщепления гласит: при скрещивании гибридов первого поколения, между собой, возникает расщепление по фенотипу в соотношении 3:1, а по генотипу 1:2:1. При скрещивании между собой черных гетерозиготных собак с генотипом Bb в помете можно ожидать рождения трех частей черных, состоящих из 1 части гомозигот BB и 2 частей гетерозигот Bb, и одной части коричневых щенков с генотипом bb. Мендель сформулировал также правило чистоты гамет, гласящее о том, гены, находящиеся в гетерозиготном состоянии не перемешиваются друг с другом, а передаются в половые клетки в неизменном виде. Определить, кто же из особей с доминантным фенотипом является гомозиготным, а кто гетерозиготным, можно только проведя, так называемое, анализирующее скрещивание с гомозиготной рецессивной формой. При таком скрещивании в случае гомозиготности исследуемой особи расщепления в потомстве не будет. В случае, же гетерозиготности — будет наблюдаться расщепление в соотношении 1:1. Еще одно правило, которое было сформулировано Менделем, носит название правила независимого расщепления аллелей. Оно состоит в том, что во втором поколении каждая пара аллелей и признаков, определяемых ими, ведет себя независимо от других пар аллелей и признаков соответственно. Для удобства анализа скрещиваний введены графические обозначения, так называемая «решетка Пеннета», в которой в верхнем ряду расписываются гаметы отца, а в левом вертикальном ряду гаметы матери. На пересечении строк и столбцов — генотипы потомков. В качестве примера приведем скрещивание гетерозиготных черных собак. Bb × Bb Племенное разведение собак Решетка Пеннета удобна тем, что она автоматически определяет все возможные генотипы и делает удобным их подсчет. В данном случае четко видно, что в потомстве данных производителей произойдет расщепление как по генотипу, так и по фенотипу. Возможное количество генотипов и фенотипов в потомстве зависит от количества пар анализируемых признаков. Нижеприведенная таблица позволяет определить числовые соотношения в потомстве при полигибридном скрещивании. Таблица 8. Числовые соотношения в потомстве при полигибридном скрещивании | |
| | | | Muftocka.
Канаровед
ПРЕДУПРЕЖДЕНИЯ : "Не говори Богам, что ты в беде. Говори беде, что ты с Богами".
Сообщения : 4330
сказали СПАСИБО : 8
Возраст : 47
Откуда : Северная Европа. Юрмала
| | Тема: Re: Генетика собак 28/2/2016, 2:39 pm | |
| Качественные и количественные признаки.Все признаки, которыми обладают живые организмы, принято делить на две категории — качественные и количественные. Качественные — признаки имеющие четко различимые формы, например, окраска или генетические аномалии, передаваемые отдельными генами. На фенотипическое проявление качественных признаков условия среды практически не влияют. Для характеристики популяции по качественным признакам используют понятия частота генов и генотипов. Однако большая часть свойств организма представлена количественными признаками. Они проявляют в основном непрерывную изменчивость и могут быть измерены — рост, длина шерсти, вес. Количественные признаки, в большей степени, чем качественные, зависят от условий среды и обусловлены многими генами, так называемыми полигенами, то есть системой неаллельных генов, одинаково влияющих на формирование данного признака. Взаимодействие таких генов в процессе формирования признака называется полимерным. Эти гены также называются аддитивными, так как их действие суммируется. Распределение их численных значений в популяции приближается к кривым нормального распределения. Их наследование можно рассматривать по схеме полигибридного скрещивания. Селекционеру приходится иметь дело в основном с непрерывной изменчивостью. Менделевский подход к изучению количественных признаков затруднен, хотя они и подчиняются тем же законам классической генетики, что и качественные. Взаимодействие неаллельных геновГены, находящиеся в разных локусах также могут влиять друг на друга. При этом можно выделить несколько типов подобного взаимодействия. Гены, не проявляющие собственного действия, но усиливающие или осласнегуркающие эффект действия других генов, называются генами-модификаторами. Изучение окраски у млекопитающих показало, что наряду с крайними формами, обладающими полным развитием пигмента или его отсутствием, наблюдается целый ряд генотипически обусловленных форм. Так, белая пятнистость у собак варьирует от нескольких белых волосков на месте первичной точки депигментации до полностью белой собаки с небольшим пучком окрашенных волос в одном из пигментных центров. В пределах генотипа, определяемого локусом белой пятнистости S, можно выделить массу переходных форм. Рис. 17. Разные варианты белой пятнистости собак    | |
| | | | Muftocka.
Канаровед
ПРЕДУПРЕЖДЕНИЯ : "Не говори Богам, что ты в беде. Говори беде, что ты с Богами".
Сообщения : 4330
сказали СПАСИБО : 8
Возраст : 47
Откуда : Северная Европа. Юрмала
| | Тема: Re: Генетика собак 28/2/2016, 2:40 pm | |
| Широкую вариабельность, обусловленную генами-модификаторами, имеют чепрачный и тигровый окрасы собак.
Если признак образуется при наличии двух пар неаллельных генов, дающих при совместном действии не тот эффект, который каждый из них обусловливает самостоятельно, то такие аллели обозначают как комплементарные (дополняющие друг друга). В качестве примера комплементарности у собак обычно приводят взаимодействие генов локусов В и Е, определяющих окраску.
Рис. 18. Комплементарное взаимодействие генов, определяющих окраску собак: соотношение — 9 черных (Ч): 3 коричневых (К): 3 рыжих (Р): 1 палевая (П)
Гены локуса В отвечают за синтез черного (В) или коричневого (b) пигмента. Гены локуса Е отвечают за распределение этих пигментов. Аллель Е способствует распространению черного или коричневого пигмента по всему телу собаки. Аллель е препятствуют их синтезу в шерстном покрове. Собаки с генотипом ее — рыжие или желтые. Черный или коричневый пигмент при этом сосредоточен только на кожных покровах морды собаки.
Формирование окраски собаки зависит от присутствия обоих пар генов. Собаки с генотипом ЕЕ или Ее — черные или коричневые в зависимости от аллелей B или b. При Е-ВВ или Е-Bb — собака черная, при E-bb — коричневая.
Собаки с генотипом ееВ- — рыжие с черным носом. Собаки генотипа bbee — обычно палевые или светло-желтые со светлым носом.
Комплементарным взаимодействием генов обусловлен особый тип паралича задних конечностей у помесей датского дога с сенбернаром. Генетический анализ проведенный Стокардом (Stockard, 1936), показал, что при чистопородном разведении как датских догов, так и сенбернаров паралич не развивается.
Аналогичное заболевание отмечалось у некоторых помесных собак-ищеек (Петухов и др., 1985).
В любой паре аллелей доминантный ген препятствует (полностью или частично) проявлению своего рецессивного партнера. Но иногда действие доминантного аллеля подавляется действием гена из другого локуса. Всемогущий ген, который блокирует действие другого гена или генов называется эпистатичным. А само явление — эпистазом. Гены, действие которых подавлено, называется гипостатичными.
Так, рецессивные гены окраски собак из локуса C не позволяют синтезироваться пигментам, определяющим окраску шерсти. Собака гомозиготная по ним — белая.
Один и тот же мутантный признак может проявляться у одних и не проявляться у других особей родственной группы. Способность данного гена проявлять себя фенотипически называется пенетрантностью. Пенетрантность определяется по проценту особей в популяции, имеющих мутантный фенотип. При полной пенетрантности (100%) мутантный ген проявляет свое действие у каждой особи. При неполной пенетрантности (меньше 100%) ген проявляется фенотипически не у всех особей.
У собак достаточно часто встречаются видоизменения хвостов в виде их укороченности, разнообразных изломов и изгибов. Можно предположить, что многообразие этого признака обусловлено его неполной пенетрантностью.
Степень пенетрантности может сильно изменяться под воздействием условий среды.
Рис. 19. Схема дигибридного расщепления с рецессивным эпистазом: вВ F 2 получено 9 черных: 3 коричневых: 4 белых собаки. Таким образом, наблюдается характерное для рецессивного эпистаза отклонение от теоретически ожидаемого расщепления 9:3:3:1
Часто особи, обладающие тем же генотипом в отношении какого-либо наследственного признака, очень сильно различаются по его экспрессивности, то есть степени проявления данного признака. Один и тот же ген у разных особей в зависимости от влияния генов-модификаторов и внешней среды может проявить себя фенотипически по-разному. Внешняя среда и гены-модификаторы могут изменить экспрессию гена, то есть выражение признака.
В отличие от пенетрантности, которая указывает, у какой доли особей в популяции проявляется данный признак, экспрессивность относится к изменчивости признака у тех особей, у которых он проявляется. Так у собак экспрессивность развития прибылых пальцев варьирует от полностью развитых пальцев на обеих задних конечностях до наличия их в зачаточном состоянии только на одной конечности. Подобная вариация экспрессивности характерна и для других наследуемых признаков, в частности и для вышеупомянутых хвостов.
Экспрессивность и пенетрантность гена в значительной степени зависят, по-видимому, от влияния генов-модификаторов и условий развития особей.
Достаточно широко распространено явление плейотропии — влияния одного гена на развитие двух и более признаков. Классический «собачий» пример плейотропного влияния — действие фактора Мерля, (локус М; окраса собак). Аллель M в гетерозиготном состоянии Мm дает пятнистость типа «арлекин» характерную для догов. Аллель M в гетерозиготе Mm в сочетании с подпалом дает «мраморный» (blue-merle) окрас, типичный для колли и шелти. В гомозиготном состоянии ММ он ведет к рождению чисто белых щенков (white-merle) со значительными аномалиями органов чувств. Такие щенки часто погибают еще до рождения, а если и рождаются живыми, то их жизнеспособность резко снижена.
Явление плейотропии объясняется тем, что гены плейотропного действия контролируют синтез ферментов, участвующих в многочисленных обменных процессах в клетке и в организме в целом и, тем самым, одновременно влияющих на проявление и развитие многих признаков.
Некоторые гены вызывают столь сильные отклонения от нормы, что снижают жизнеспособность организма или даже приводят его к гибели. Такие гены называют летальными, то есть смертоносными, или сублетальными — снижающими жизнеспособность. В большинстве случаев летальные гены полностью рецессивны, поэтому гетерозиготные носители этих генов фенотипически совершенно неотличимы от нормальных особей. В гомозиготном состоянии такие гены могут нарушать нормальный ход развития эмбриона на любых стадиях. О возможности наличия летальных генов можно косвенно судить по снижению средней численности пометов или по выпадению некоторой ожидаемой части фенотипов в расщеплении.
Так в случае скрещивания упоминавшихся выше черно-мраморных собак, гетерозиготных по фактору Мерля получается вместо ожидаемого соотношения 3:1, получается 2:1 т. е. 2 мраморных и 1 черная собаки Mm × Mm = ММ: 2 Mm: mm, где ММ белая нежизнеспособная собака. Белые щенки чаще не рождаются вовсе, так как погибают задолго до рождения.
Рис. 20. Наследование мраморности типа «арлекин» у догов. Ген М h (фактор Мерля) — доминантный с рецессивным летальным действием: 1 — скрещивание мраморных догов между собой; 2 — анализирующее скрещивание
Фактор Мерля относится к категории доминантных летальных генов, которых значительно меньше, чем рецессивных. При желании его носителей легко изъять из разведения, так как они имеют характерный фенотип. Одни летальные гены обуславливают крупные аномалии, другие — нарушения физиологических процессов. Пути губительного воздействия большинства летальных генов не ясны. Таких генов может существовать сколь угодно много. Показано, что каждый человек является носителем в среднем 4–9 «вредных» или летальных генов. Аналогичных результатов можно ждать и по собакам. Известны летальные гены, которые при проявлении в эмбриональном состоянии, опасны и для жизни беременной суки, например, при наследственной контрактуре мышц плода, вследствие чего сука не может разродиться.
Взаимодействие генов, когда при их сочетании в одном организме развивается совершенно новая форма признака называется новообразованием.
Иногда новообразования приводят к появлению признаков дикого фенотипа. В этом случае их называют атавизмами, то есть возвратом к предковой форме или возвратом в дикий тип.
Частичные возвраты в дикий тип возможны при скрещивании двух особей одной породы, если эти производители происходят из удаленных друг от друга неродственных популяций. По-видимому, подобным образом можно объяснить и большое сходство между дворнягами, обитающими в самых разных местах.
Признаки, сцепленные с полом
Признаками, сцепленными с полом называют такие, которые формируются под воздействием генов, локализованных в Х-хромосоме. Наиболее характерным примером наследования признака сцепленного с полом является наследование гемофилии собак. У собак больных гемофилией, в крови не хватает фактора, который, взаимодействуя с кровяными пластинками (тромбоцитами), ускоряет превращение протромбина в тромбин. Гемофилия собак аналогична гемофилии человека и также обусловлена сцепленным с полом рецессивным геном. Ген, определяющий развитие гемофилии находится в Х-хромосоме и является рецессивным по отношению в нормальному аллелю. Следовательно, гемофилия проявляется только у гомозиготных самок (несущих этот ген в обеих Х-хромосомах) и гемизиготных самцов, несущих ген гемофилии в Х-хромосоме. Щенки-гемофилики обычно погибают в раннем возрасте от наружных или внутренних кровотечений. Сохранить до половозрелого состояния такого самца возможно только при постоянном введении специфических препаратов. Самки же неминуемо гибнут не позже первой течки. Гетерозиготные самки внешне абсолютно нормальны и плодовиты. Однако половина их детенышей мужского пола страдает гемофилией и половина детенышей-самок оказывается гетерозиготными по этому гену.
Родители:
сука-носительница гена гемофилии
ХHХh
кобель нормальный
XHY
— половая хромосома, несущая нормальный аллель
XH
Xh — половая хромосома, несущая ген гемофилии
К признакам сцепленным с полом относятся также врожденный гипотрихоз, отмеченный у такс и карликовых пуделей; мышечная дистрофия у ретриверов; синдром пошатывания, связанный с гипомиелинизацией и обнаруженный у чау-чау и ряда других пород; подвывих запястья; а также диафрагмальная грыжа, описанная у золотистых ретриверов. | |
| | | | Muftocka.
Канаровед
ПРЕДУПРЕЖДЕНИЯ : "Не говори Богам, что ты в беде. Говори беде, что ты с Богами".
Сообщения : 4330
сказали СПАСИБО : 8
Возраст : 47
Откуда : Северная Европа. Юрмала
| | Тема: Re: Генетика собак 28/2/2016, 2:42 pm | |
| | |
| | | | Muftocka.
Канаровед
ПРЕДУПРЕЖДЕНИЯ : "Не говори Богам, что ты в беде. Говори беде, что ты с Богами".
Сообщения : 4330
сказали СПАСИБО : 8
Возраст : 47
Откуда : Северная Европа. Юрмала
| | Тема: Re: Генетика собак 28/2/2016, 2:44 pm | |
| У собак нашли ген альбинизма, схожий с человеческимПейдж Винклер, докторант в College of Veterinary Medicine, и соруководитель исследования Джошуа Барто, доцент Department of Small Animal Clinical Sciences, обнаружили мутантный ген у доберманов-альбиносов, похожий на одну из форм альбинизма у человека.  «Мы обнаружили мутацию, которая приводит к исчезновению белка, необходимого для клетки, чтобы стать пигментированной», сказал Винклер. «Некоторые дефекты этого же гена вызывают состояние, которое называется глазокожный альбинизм у человека.» Этот тип альбинизма имеет определенные характеристики, которые отмечены у людей и собак. «У доберманов-альбиносов вы увидите белую или сильно осветленную шерсть, розовый нос и губы с бледную радужку глаз», — сказал Винклер. «Эти черты очень схожи с характеристиками людей с глазокожным альбинизмом, имеющих светло-пигментированные кожу и волосы, а также светлые глаза и нарушения зрения.» 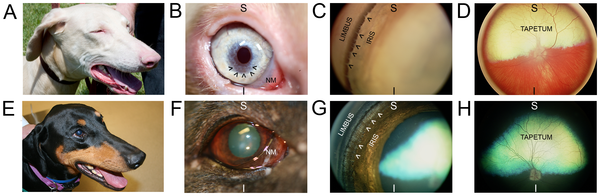 Собаки этой породы и люди также имеют высокую чувствительность кожи к солнечным лучам, что приводит к повышенному риску возникновения опухолей кожи. «Мы знали, что у доберманов-альбиносов обычно развиваются эти типы опухолей, так же, как и у людей, и мы обнаружили значительное увеличение риска развития меланомы у собак-альбиносов.» Барто и Винклер изучили по 20 собак с нормальной пигментацией и альбиносов и обнаружили, что более половины альбиносов имеют по крайней мере одну опухоль, в то время как только одна из собак с нормальной пигментацией имела опухоль. Результаты их исследования могут иметь большое значение для заводчиков доберманов по всему миру. «Поскольку доберманы могут нести дефектный ген, но не показывать никаких признаков патологии, это создает серьезные проблемы для заводчиков», сказал Барто. «Но теперь, когда мы определили мутацию, мы можем начать тестировать на носительство этого гена». Результаты исследования опубликованы в PLoS ONE . Источник: http://cynology.bestforums.org/ http://www.redorbit.com/news/science/1113144997/humans-dogs-share-similar-albino-gene-051314/ | |
| | | | Muftocka.
Канаровед
ПРЕДУПРЕЖДЕНИЯ : "Не говори Богам, что ты в беде. Говори беде, что ты с Богами".
Сообщения : 4330
сказали СПАСИБО : 8
Возраст : 47
Откуда : Северная Европа. Юрмала
| | Тема: Re: Генетика собак 28/2/2016, 2:48 pm | |
| Скрытая опасность ЛайнбридигнаНедавно было опубликовано еще одно исследование, описывающее рецессивную мутацию, которая появилась из ниоткуда, чтобы разрушить жизни некоторых щенков лабрадор ретривера с врожденным миастеническим синдромом — нервно-мышечным заболеванием, которое приводит к слабости мышц, нарушая передачу химических сигналов, вызывающих их сокращение. Это произошло в помете от пары родителей, имеющих двух общих предков. Исследователи также изучили родственников этих собак и обнаружили, что 16 из 58 были носителями этой мутации, т.е. почти 30%. В то время как 288 не родственных им лабрадоров не имели подобной мутации.  Авторы пишут: «лайнбридинг в этой семье лабрадор ретриверов делает возможным то, что отец и мать унаследовали мутацию от общего предка, и что заболевшие щенки гомозиготны по этому сегменту хромосомы»… «практика лайнбридинга ускоряет появление рецессивных заболеваний в чистокровной популяции». Давайте разберемся. Это особенное семейство (линия) несет мутацию, которая вызывает серьезное генетическое заболевание, в данном случае — нарушает нервно-мышечную связь у собак, которые получают две копии дефектного гена. Собаки, имеющие только одну копию, по всей видимости здоровы. Повязав двух собак с общими предками, которые, как выяснилось, были носителями, получили щенков с патологией. Таким образом знание кровей и линий никак не помогло избежать или узнать об этой мутации, скрытой в этой линии собак. Коэффициент инбридинга мог отразить вероятность получение двух копий гена с обеих сторон родословной. Чем выше КИ, тем больше риск того, что это произойдет. Все собаки имеют мутации, о которых заводчик не имеет возможности узнать. Если говорить об «ответственных заводчиках», то перед вязкой они бы сделали все доступные ДНК тесты для наследственных заболеваний, известных в этой породе. Но даже это и знание линий и кровей собак не дает возможность избежать подобных ситуаций. «Знание кровей» хороший совет для начинающих заводчиков, однако это не дает вам знание всего, что может произойти. К тому же ДНК-тесты не могут решить проблему наследственных заболеваний чистокровных собак. Единственный способ «контролировать неизвестность» — это селекция с минимальным риском возможности встречи двух носителей скрытой мутации. Поэтому заявления заводчиков о том, что они «знают свои линии, и поэтому могут применять лайнбридинг без последствий» не имеют под собой никаких оснований. Я не знаю, действительно ли такие заводчики верят в видимость того, что в их линиях нет никаких проблем, или, возможно, это более серьезный миф, что можно избежать проблем инбридинга, «зная свои линии». Это всего лишь вопрос времени, когда такие «подводные мины» дадут о себе знать. Подобные заверения о «знании» и «опыте», которые почему-то дают им право размножать близкородсвенных животных «без последствий», хотя на самом деле они участвуют в игре под названием рулетка, в которой рано или поздно проигравшим станет щенок и семья с разбитым сердцем, которая его получит. Rinz CJ, J Levine, KM Minor, HD Humphries, R Lara, AN Starr-Moss, LT Guo, DC Williams, GD Shelton, & LA Clark 2014 A COLQ Missense mutation in Labrador Retrievers having congenital myasthenic syndrome. PLoS One 9(8): e106425. http://cynology.bestforums.org/ Автор: Carol Beuchat (переведено с разрешения автора). Оригинал: The fiction of «knowing your lines» | |
| | | | Muftocka.
Канаровед
ПРЕДУПРЕЖДЕНИЯ : "Не говори Богам, что ты в беде. Говори беде, что ты с Богами".
Сообщения : 4330
сказали СПАСИБО : 8
Возраст : 47
Откуда : Северная Европа. Юрмала
| | Тема: Re: Генетика собак 28/2/2016, 2:56 pm | |
| Цена инбридинга: Даже 100 лет назад достоинства и недостатки инбридинга были очевидны, хотя механизмы этого явления не были до конца понятны, опытные заводчики могли найти компромиссы. Понятие «коэффициента инбридинга» [Райта] (КИ) было специально сформулировано, чтобы использовать в качестве инструмента для количественной оценки издержек и выгоды в селекционной работе. Перечислим их: ПРЕИМУЩЕСТВА - НЕДОСТАТКИ — увеличение однородности — снижение плодовитости — увеличение препотентности (способности передавать характерные черты потомству) — снижение приспособленности — «фиксация» желаемых черт и породного типа — врожденные дефекты — уменьшаются размеры — снижение количества щенков в помете — медленный рост — повышение смертности щенков — сокращение продолжительности жизни — увеличение генетических заболеваний — уменьшение «генетического потенциала» (способности улучшать черты) Почему инбридинг увеличивает однородность и препотентность? Инбридинг увеличивает гомозиготность. Под этим мы подразумеваем, что если есть два (или более) аллеля в популяции для гена, частота встречаемости АА и аа увеличится, тогда как Аа и аА сократится. Если собака является гомозиготной по АА, то она может передавать только аллель А своим потомкам. Чем больше степень гомозиготности генома животного, тем более однородным будет его потомство, потому что они все будут иметь одинаковые аллели для многих генов этого родителя. Если вы стараетесь разводить по определенному признаку — то это дает огромное преимущество. Вариабельность среди однопометников будет снижена, и инбредный родитель будет «доминантным», т.е. дающий потомство с узнаваемыми чертами. И все было бы хорошо, только природе не особо нравится гомозиготность. В сотнях исследований по различным видам растений и животных (в том числе домашних животных) имеются неопровержимые доказательства того, что за гомозиготность нужно платить. Райт отметил, что «снижение жизнеспособности при применении инбридинга в ранее хаотичной популяции может быть прямо пропорционально увеличению доли гомозиготности» и «что касается других последствий инбридинга: «фиксация» черт и повышение препотентности, также, конечно, находится в прямой зависимости от процента гомозиготности». Это отображено на рисунках. «Приспособленность» является мерой способности животного производить потомство, которая может способствовать передаче генов последующим поколениям. Производство потомства, которое само не дает потомства, имеет приспособленность равную нулю. Такое может произойти, если потомство бесплодно, или умерло сразу после рождения, или было слишком медленным или глупым, чтобы убежать от хищника — не имеет значения, важно лишь — останутся ли гены данного животного в генофонде через поколение.  Когда КИ = 0, приспособленность находится на максимуме, а препотенция и однородность на минимуме. При увеличении инбридинга вариабельность у потомков снижается, но также снижается и приспособленность. Инбридинг снижает приспособленность (красная линия), и уменьшение приспособленности пропорционально степени инбридинга. Результат снижения приспособленности называется «инбредная депрессия», которая представляет собой весь спектр проявлений снижения приспособленности. Если вы начнете с «беспородного» растения (имеющего высокий гетерозис) и произведете самоопыление (самооплодотворение, без применения партнера для размножения), всхожесть семян будет снижена. При продолжении самоопыления в последующих поколениях снижение всхожести семян будет пропорционально коэффициенту инбридинга и величине гомозиготности (синяя линия).  Это очень удобные соотношения. Издержки и выгоды инбридинга прямо пропорциональны КИ. Это означает, что заводчик может выбрать уровень инбридинга, который имеет хороший баланс между его издержками и выгодами. Инбредная депрессия у собак Существуют ли данные, отображающие отрицательный эффект инбредной депрессии у собак? Вот некоторые данные о численности пометов 6 пород собак из документов Шведского кеннел клуба. Первое, о чем необходимо предупредить, то, что они, вероятно, недооценивают истинный КИ (рассматривая до 10 поколений), особенно для пород, популярных в последнее время (таких, как голдены и лабрадоры). Поэтому размещение линий на оси X относительно друг друга, вероятно, будет не совсем 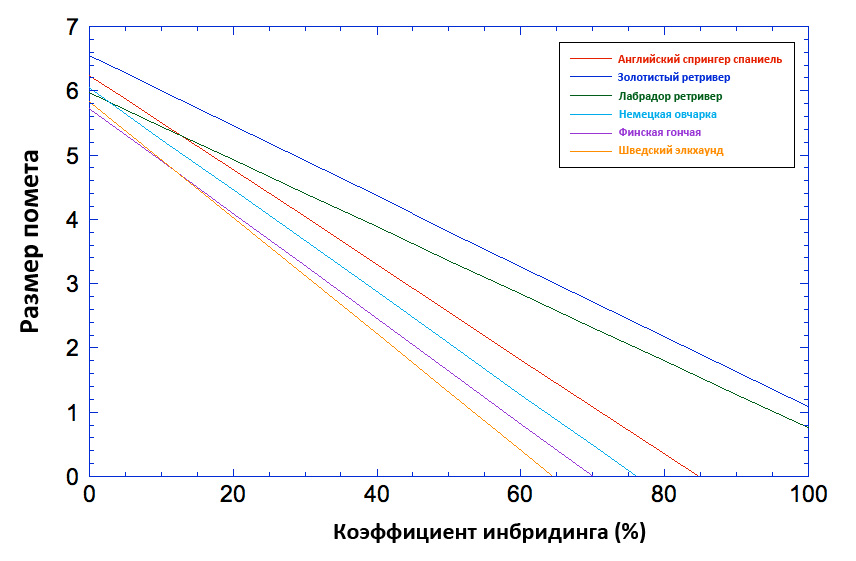 Для наших целей, однако, это не имеет особого значения. То, на что нам следует обратить внимание — это эффект КИ на размер помета, и как отметил Райт и другие исследование, существует линейная зависимость сокращения размера помета и увеличения инбридинга. Эти линии наглядно демонстрируют нам отношение издержек и выгод. Для этих пород это отношение равно 0,1, что означает, что увеличение инбридинга на 10% снижает размер помета примерно на 1. Если нормальный размер помета составляет около 6 щенков, при КИ 30% — что не редкость во многих породах — размер помета уменьшится на половину. Т.е. вдвое меньше щенков, из которых можно выбрать лучших, и к тому же эти щенки будут менее приспособленными, согласно Райту, они будут меньше, менее энергичные, будут иметь больше врожденных дефектов и более высокую смертность, будут расти медленнее, иметь более короткую продолжительность жизни, и, конечно, увеличится число генетических болезней, вызываемых рецессивными мутациями. Как инбредная депрессия отражается на потомках? Это отчетливо прослеживается на смертности щенков. Вот данные по биглям, у которых при КИ менее 25% смертность щенков до 10 дневного возраста составила более 20%. При увеличении КИ более 25% увеличивается и смертность, 30% для собак с КИ 25-50%, и выше, когда КИ еще увеличивается. У пуделей с КИ менее 6% продолжительность жизни на 4 года больше, чем с более высоким КИ, а риск заворота желудка примерно пропорционален увеличению КИ, т.е. увеличение КИ на 10% увеличивает риск развития заворота примерно на то же число. 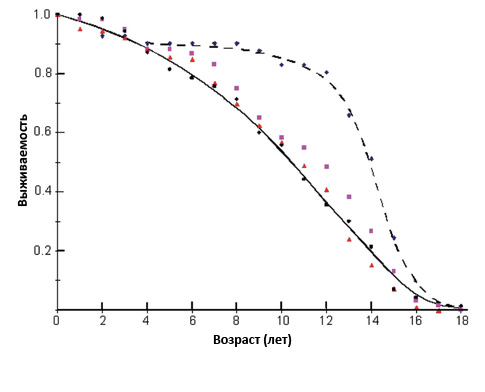 Продолжительность жизни стандартных пуделей при различных уровнях инбридинга. Голубые ромбы: <6,25% (N=39), розовые квадраты: 6,25-12,5% (N=65); красные треугольники: 12,5-25% (N=141); черные кружки: >25% (N=71). Непрерывная линия отображает группу >25%. 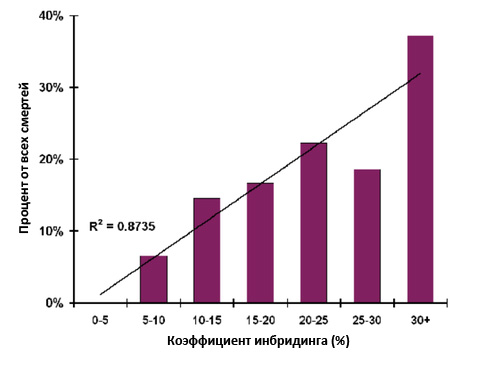 Зависимость КИ и данных о смерти от заворота желудка. У бернских зенненхундов при каждом увеличении КИ на 10% сокращает продолжительность жизни на 200 дней. Для собак с КИ 30% продолжительность жизни снижается почти на 2 года. 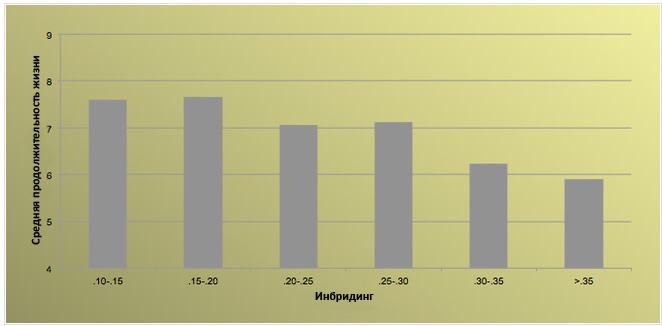 Так сколько инбридинга слишком много? Теперь этот вопрос на миллион долларов. Если бы инбридинг нес только хорошее, селекционеры бы использовали только его и животные процветали бы. Но заводчикам необходимо контролировать инбридинг по причинам, описанным выше. Итак, как же оценить издержки и выгоды? Правило для устойчивого развития диких и домашних животных — это держать инбридинг ниже 5%, и использовать 10% как верхний предел для популяций. Там могут встречаться животные, которые имеют КИ гораздо выше (например, для сохранения ценных качеств и скрещивания в пределах линии), но когда среднее количество в популяции выше 10%, начинаются проблемы. Сокращается плодовитость, пометы становятся меньше, размер животных уменьшается, увеличивается риск развития генетических заболеваний, сокращается продолжительность жизни, снижается приспособленность. Получается отрицательная обратная связь, что называется «вихрь вымирания» (extinction vortex), и как только популяция вступает на этот путь, становится очень сложно остановить этот процесс. Отметим еще раз, что порог для запуска «вихря» составляет КИ в 10%. У большинства пород домашнего скота (коров и т.д.), КИ сохраняется ниже этого предела. При разведении служебных рабочих собак также придерживаются КИ менее 10%. Но в среде породистых собак дело обстоит иначе, КИ (рассчитываемый назад к основателю) может быть сложно определим. И свидетельства инбредной депрессии и других последствий инбридинга очень очевидны — пометы с единственным щенком, высокие темпы развития рака у молодых собак, аллергии и многие другие показатели, которые кажутся «нормой» в современном разведении.  Инбридинг может быть использован для получения исключительных животных, но цена этого может быть очень велика! Автор: Carol Beuchat (перевод осуществлен с разрешения автора). Оригинал: The Costs and Benefits of Inbreeding и Inbreeding Effects http://cynology.bestforums.org/ | |
| | | | Muftocka.
Канаровед
ПРЕДУПРЕЖДЕНИЯ : "Не говори Богам, что ты в беде. Говори беде, что ты с Богами".
Сообщения : 4330
сказали СПАСИБО : 8
Возраст : 47
Откуда : Северная Европа. Юрмала
| | Тема: Re: Генетика собак 5/3/2016, 2:52 pm | |
| Ой, мамочки! Думала эту темку продолжить, но решила все-таки отдельно. А то вообще запутаемся  | |
| | | | | | Генетика собак | |
|
Похожие темы | |
|
| | Права доступа к этому форуму: | Вы не можете отвечать на сообщения
| |
| |
| |





